


Abstract
Background: Lung cancer cells often express vimentin. However, the function of vimentin in lung cancer cells has not been fully evaluated. Materials and Methods: We evaluated the association between vimentin expression in resected non-small cell lung cancer (NSCLC) specimens and prognosis. Short-interfering RNA targeting vimentin and establishment of an invasive cell line by repeated selection of invasive cells using a Matrigel membrane invasion chamber system (MICS) were performed. MICS was used to reveal the relationship between invasiveness and vimentin. Results: Vimentin positivity was significantly associated with a poor prognosis and was significantly lower in squamous cell carcinoma than in adenocarcinoma. In in vitro experiments, silencing of vimentin reduced invasiveness. Highly invasive cell lines exhibited higher expression of vimentin than did parental cells, and invasive ability was reduced by knockdown of vimentin. Conclusion: Vimentin expression is associated with prognosis via alteration of the invasive ability of NSCLC cells.
Vimentin, a 57-kDa protein, is one of the most widely expressed proteins of the type III intermediate filament protein family and normally expressed in cells of mesenchymal origin (1). Many studies have evaluated the relationship between vimentin and cancer. Vimentin has been detected in poorly differentiated prostate cancer, and down-regulation of vimentin reduced the invasive ability of these cancer cells (2). Vimentin expression has most often been associated with the invasive ability of gastric cancer and was suggested to play an important role in metastasis (3). A non-invasive breast cancer cell line acquired increased motility and invasiveness upon vimentin overexpression, and these characteristics were down-regulated by silencing of vimentin in an invasive tumor cell line that constitutively expressed vimentin (4). Some studies have also investigated the significance of vimentin in lung cancer and found that vimentin expression was associated with a poor prognosis (5) and a high frequency of metastases in patients with resected non-small cell lung cancer (NSCLC) (6). However, the mechanisms by which vimentin affects the prognosis of NSCLC have not been determined. Therefore, we studied the relationship between vimentin and prognosis, and the role of vimentin in lung cancer invasion.
Materials and Methods
Patients. NSCLC tissue samples were obtained from 108 patients with primary lung cancer who underwent surgical resection from 1995 to 1999 at Kagawa University Hospital. Written informed consent statements were obtained from all patients before surgery. Formalin-fixed, paraffin-embedded primary lung cancer specimens were acquired from the Department of Diagnostic Pathology, Kagawa University, under approval from the Institutional Review Board of Kagawa University and principles of the Declaration of Helsinki. The tumors were staged according to the sixth edition of the TNM International Union Against Cancer/American Joint Committee on Cancer classification.
Vimentin immunostaining. Tissue sections (4 μm) were created from formalin-fixed, paraffin-embedded blocks of NSCLC specimens. Serial tissue sections were immunohistochemically examined using antibodies to vimentin (Ref M0725, dilution 1:200, clone V9; Dako, Glostrup, Denmark) using a Ventana BenchMark XT autostainer (Ventana Medical Systems, Inc., Tucson, AZ, USA). All areas of the tumor were evaluated, and the percentage of tumor cells that expressed vimentin at any intensity level was calculated. We defined a positive case as the presence of >10% vimentin-expressing tumor cells.
Cells and reagents. A549, HI1017, and RERF-LC-OK (derived from adenocarcinoma of the lung) cell lines were obtained from the Japan Cancer Research Bank (Tokyo, Japan). Cells were cultured in RPMI-1640 supplemented with 10% fetal bovine serum (FBS). The antibodies used were AC-15 (β-actin; Sigma-Aldrich, St. Louis, MO, USA), D21H3 (vimentin; Cell Signaling Technology, Danvers, MA, USA), and anti-mouse immunoglobulin G conjugated with horseradish peroxidase (Santa Cruz Biotechnology, Santa Cruz, CA, USA). Short interfering RNAs (siRNAs) targeting vimentin and control siRNA were purchased from Sigma-Aldrich. Short hairpin RNAs (shRNAs) targeting vimentin and control shRNA were purchased from Takara Bio (Shiga, Japan).
Invasion assays. An in vitro invasion assay was performed as previously described (7). Briefly, 1.0×105 cells suspended in RPMI-1640 without FBS were seeded into the upper wells of 24-well Matrigel-coated invasion chambers (Becton Dickinson, Franklin Lakes, NJ, USA). RPMI-1640 supplemented with 10% FBS was added to the lower wells as a chemoattractant. After incubation for 24 h, cells that invaded through the membrane were fixed in methanol, stained with Diff-Quik stain, and microscopically counted.
Establishment of invasive cells. Sub-populations from A549 cancer cell lines were selected according to their differential invasiveness using Matrigel-coated invasion chambers as previously described (7). Similar to invasion assays, cells were seeded into the upper wells. Following incubation for 12 to 72 h at 37°C, the inserts were removed. The cells that migrated through the Matrigel membranes and attached to the lower wells were allowed to proliferate. This selection of invasive cells was repeated 9 and 18 times to establish highly invasive sublines. Parental cells were designated A549-0, and invasive cells that migrated through the Matrigel 9 and 18 times were designated A549-9 and A549-18, respectively. To assess the invasive ability, highly invasive A549-9 and A549-18 cells were incubated for 8 h, which was shorter than the regular assay.
RNA interference. RNA interference was performed to selectively silence vimentin. Cells were seeded in 6-cm dishes at a density of 4×105 cells/dish. The next day (day 0), cells were transfected with siRNA (final concentration, 50 nM) in Opti-MEM I (Invitrogen, Carlsbad, CA, USA) using Lipofectamine 2000 (Invitrogen). After 8 h of incubation, the media were replaced with RPMI-1640 supplemented with 10% FBS. Cell lysates were extracted on day 4, and the efficacy of RNAi was assessed by immunoblots.
To establish stable transfectant, cells were seeded in 6-cm dishes at a density of 4×105 cells/dish. The next day (day 0), cells were transfected with vimentin shRNA or control shRNA in Opti-MEM I using Lipofectamine 2000 and PLUS reagent (Invitrogen). After 8 h of incubation, the media were replaced with RPMI-1640 supplemented with 10% FBS. Cells were selected in the presence of Puromycin (Invitrogen) for stable expression of shRNA. We established two types of each cell line (shVIM1, shVIM2, Cont1, and Cont2).
Immunoblots. Immunoblots were performed as previously described (8). Briefly, total cell lysate from cancer cells was prepared by homogenizing the cells in 2× sodium dodecyl sulfate (SDS) sample buffer; this mixture was boiled and subjected to SDS-polyacrylamide (10%) gel electrophoresis followed by immunoblotting using AC-15 and D21H3. The intensity of positive signals in the immunoblot analyses was imported by Image Reader LAS-1000 Plus (Fuji Photo Film Co. Ltd., Tokyo, Japan) and quantified densitometrically using NIH ImageJ 1.45s (NIH, Bethesda, MD, USA).
Statistical analysis. Quantitative variables are described as mean±standard deviation. Univariate analysis was performed to identify factors associated with the occurrence of events (death) using the Kaplan–Meier method and compared with the log-rank test. A value of p<0.05 was considered statistically significant. Survival data were evaluated using a multivariate Cox proportional hazard model. All analyses were performed using Excel Statistics version 2012 (Social Survey Research Information, Tokyo, Japan).
Results
Expression of vimentin in clinical samples and relationship to prognosis. The characteristics of the 108 patients are shown in Table I. Immunohistochemical staining was used to analyze vimentin expression in tumor specimens. Representative immunohistochemical stain findings are shown in Figure 1. Vimentin expression was positive in 40 out of 108 patients (Table I). Vimentin was found to be significantly associated with a poor prognosis (p=0.038) (Figure 2). Although we analyzed clinical parameters including age, sex, smoking status, differentiation, and lymph node status, we found no significant association with vimentin positivity. However, the vimentin-positive rate was significantly higher in adenocarcinoma than in squamous cell carcinoma (56.0% vs. 22.4%, respectively; p<0.01). Multivariate analysis by Cox proportional hazards model showed that pathological stage (IIA or more), squamous cell carcinoma histology and vimentin expression were identified as factors predictive of poor prognosis (Table II).
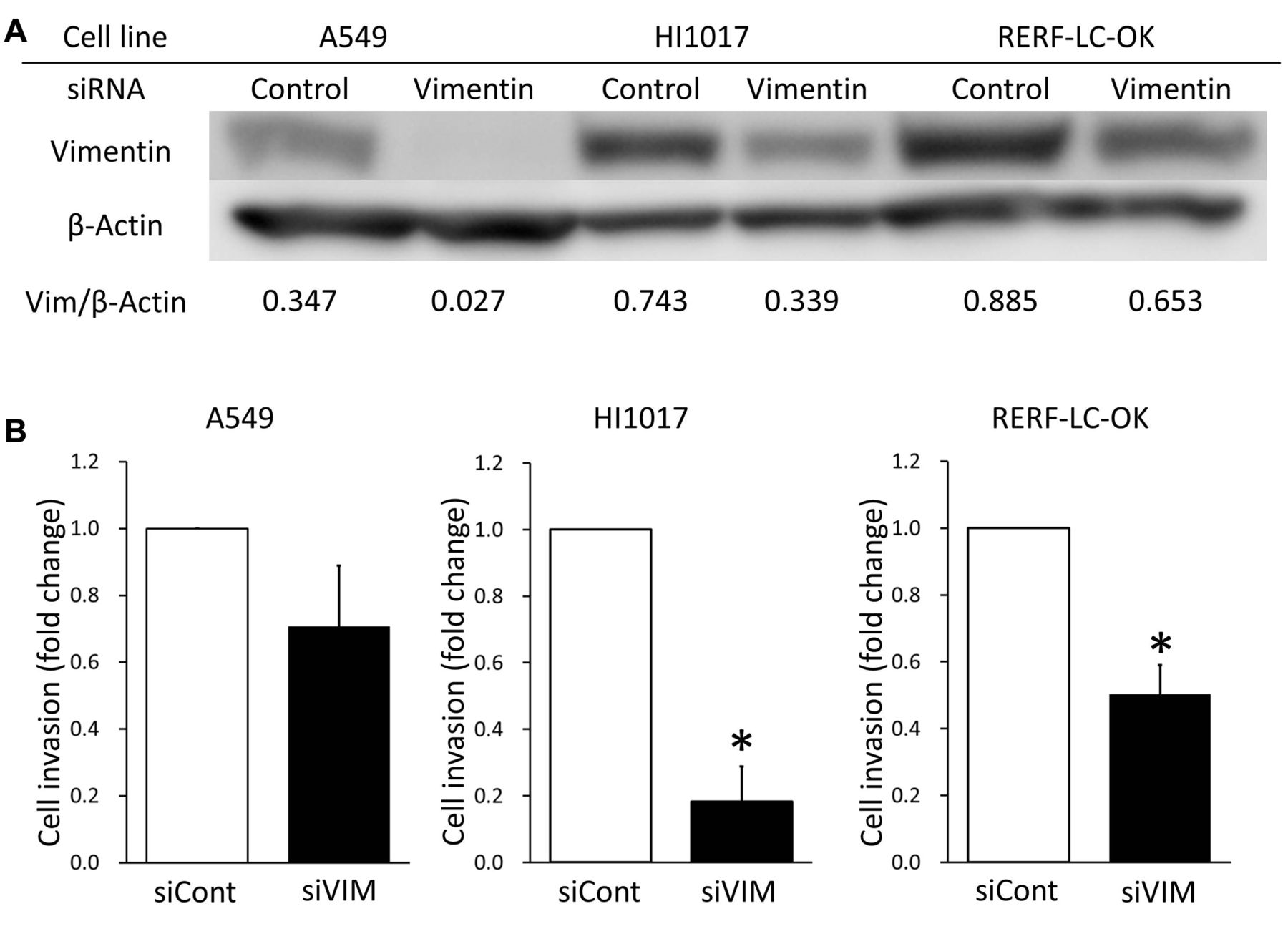
Vimentin-silenced cells have low invasive ability. To investigate the role of vimentin in invasiveness, we performed RNAi targeting vimentin in three NSCLC cell lines. Immunoblots showed that siRNA targeting vimentin reduced vimentin expression in A549, HI1017, and RERF-LC-OK cells (Figure 3). The invasion assay revealed that vimentin-knockdown HI1017 and RERF-LC-OK cells led to their having significantly less invasive ability than cells transfected with control siRNA (p<0.05) (Figure 3). However, no significant decrease in invasiveness was observed in A549 cells. Furthermore, we performed stable transfection of shRNA to HI1017 cells. Immunoblots showed that shRNA reduced vimentin expression, and the invasion assay revealed a decrease in invasive ability (Figure 4).
Vimentin expression in invasive sub-populations of A549 cells. Invasive subpopulations of A549 cells were established by repeated selection of invasive cells using Matrigel-coated membranes. A549 cells that migrated through the Matrigel 9 and 18 times (A549-9 and A549-18, respectively) showed a greater invasive ability. Eight-hour incubation was enough for A549-9 and A549-18 cells to migrate across the membrane, whereas only a few A549-0 cells were able to migrate (Figure 5). Interestingly, vimentin expression was higher in invasive cells than in parental A549 cells (Figure 5). To further investigate the function of vimentin in the invasiveness of highly invasive A549 cells, we performed RNAi to knock-down vimentin. The invasion assay revealed that vimentin-knockdown of A549-18 cells led to a significantly lower invasive ability than did the control (Figure 5).

Immunohistochemical staining for vimentin in resected specimens of non-small cell lung cancer. Examples of vimentin-positive (A) and vimentin-negative (B) specimens.

Kaplan–Meier curves for overall survival. The curves are based on the vimentin expression status (positive and negative, p=0.038).

Invasive abilities of three non-small cell lung cancer cells after transfection with siRNA targeting vimentin. A549, HI1017, and RERF-LC-OK cells were transfected with siRNA targeting vimentin (siVIM) or control siRNA (siCont). A: The efficacy of siRNA was evaluated by immunoblotting. B: Invasive ability of A549, HI1017, and RERF-LC-OK cells were assessed by a Matrigel invasion assay. Data presented are the means of fold changes±SD of three independent experiments. *p<0.05 Compared to control siRNA.
Discussion
This study demonstrated that i) positive vimentin expression was associated with a poor prognosis in patients with surgically resected NSCLC, ii) the vimentin-positive rate was lower in squamous cell carcinomas than in adenocarcinomas, iii) established invasive A549 cells expressed more vimentin than did parent cells, and iv) suppression of vimentin reduced the invasive ability of all three tested NSCLC cell lines. The latter three findings were shown for the first time in the current study. Furthermore, these data suggest that vimentin is associated with prognosis via regulation of the invasive ability of lung cancer cells, at least in part.
In the present study, vimentin expression was positive in 37.0% of 108 NSCLC cases with a cut-off point of 10% for vimentin positivity. In previous studies, vimentin expression was detected in 26.1% of NSCLC specimens (5). Other studies reported different cut-off points of vimentin positivity. Vimentin expression was detected in 29.3%, 53.3%, and 17.5% of cases with a cut-off point of 10%, 25%, and 50%, respectively (9-11). Our results can be considered in line with these reports.
Several studies have reported that vimentin expression is associated with the occurrence of metastasis (6) and a poor prognosis in patients with lung cancer (5, 9). Consistent with these reports, a similar prognosis was observed in our study. In general, lymph node status is an important prognostic factor in NSCLC. However, no significant association between the lymph node status and vimentin positivity in the primary tumor was observed in the current study, nor in a previous study (6). Conversely, vimentin expression was associated with the occurrence of distant metastasis after surgery (6). These findings may suggest that vimentin expression is associated with hematogenous, but not lymphogenous metastasis. Cancer cells expressing vimentin might tend to invade vessels. Another interesting finding of the current study is that vimentin expression was higher in adenocarcinomas than in squamous cell carcinomas. Distant metastases are more often recognized in adenocarcinomas than in squamous cell carcinomas (12). A difference in vimentin expression might cause a difference in the distant metastasis status between adenocarcinomas and squamous cell carcinomas.
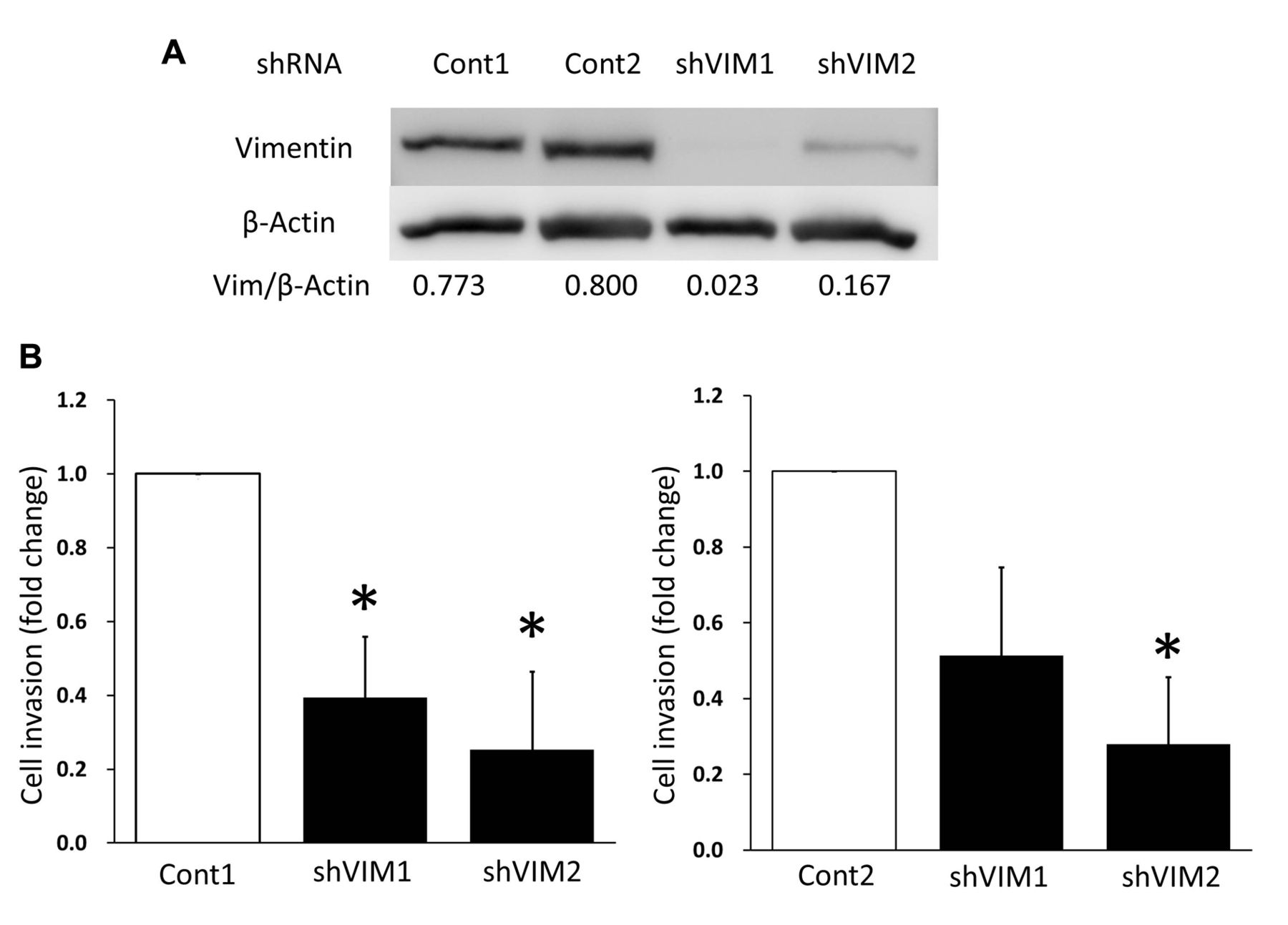
Invasiveness of HI1017 cells after silencing of vimentin with shRNA. HI1017 was transfected with shRNA targeting vimentin (shVIM1, shVIM2) or control (shCont). A: The efficacy of shRNA was evaluated by immunoblotting. B: Invasive ability was assessed by a Matrigel invasion assay. Data presented are the means of fold changes of three independent experiments. *p<0.05 Compared to shCont1 or shCont2.
Patients' characteristics (n=108).
Association between survival and clinicopathological characteristics.

Vimentin expression in invasive sub-population of A549 cells and the effect of silencing of vimentin. Invasive sub-populations were selected by repeated passages through Matrigel-coated membranes 9 (A549-9) and 18 times (A549-18). A: Expression of vimentin in A549-0, -9, and -18 cells was evaluated by immunoblots. B: Invasive ability of A549-0, -9, and-18 as assessed by a Matrigel invasion assay. Data presented are the means of fold changes±SD of three independent experiments. *p<0.05 Compared with A549-0. An invasive sub-population of A549 cells (A549-18) was transfected with siRNAs. C: The efficacy of siRNA was evaluated by immunoblotting. D: Invasive ability of A549-18 was assessed by invasion assay after transfection of siRNA targeting vimentin (siVIM) or control (siCont). Data presented are the means of fold changes±SD from three independent experiments. *p<0.05 Compared to control siRNA.
A549 cells in which vimentin was silenced did not exhibit significantly less invasive ability than control cells. As shown in Figure 3, the expression of vimentin in A549 cells was lower than in the other cell lines. Various factors have been reported to be associated with the invasiveness of the tumor cells. Therefore, we consider that invasiveness of A549 cells is defined by many factors besides vimentin and those have an impact on invasiveness more than vimentin itself does.
The findings of the current study have deepened the understanding of the association between cancer invasion and vimentin expression. We demonstrated that an invasive sub-population of lung cancer cells has higher expression of vimentin and that silencing of vimentin is associated with low invasiveness in three cell lines. These results suggest that high expression of vimentin is directly associated with the invasive phenotype of lung cancer. Some reports support a role of vimentin in invasiveness. For example, silencing of vimentin causes dysfunctional polarization of the Golgi apparatus during cell migration via a decrease in scribbled planar cell polarity protein, that is associated with cell polarization in migrating epithelial Madin–Darby canine kidney cells (13). In addition, expression of AXL receptor tyrosine kinase, a protein associated with metastasis (14), is reduced by vimentin knockdown (15). Furthermore, vimentin is a known marker of epithelial-to-mesenchymal transition (EMT), that is characterized by diminished epithelial characteristics and enhanced mesenchymal characteristics (16). Vimentin is considered one of the best indicators of EMT in carcinomas (17). Evidence for the role of EMT in tumor progression has been increasing. Thus, vimentin should be associated with the malignant phenotype in conjunction with various molecules or EMT.
Our study has several limitations. Firstly, we compared the vimentin-negative and -positive groups. There is currently no established standard cut-off for vimentin. However, we determined the cut-off as 10% based on a previous reference (6) in which the cut-off was based on the result that provided the greatest statistical significance and lowest hazard ratio. Secondly, we could not determine the effect of overexpression of vimentin on invasive ability. Although we tried to force expression in some cell lines, no difference was revealed because of minimal vimentin overexpression compared to the greater endogenous level.
In summary, vimentin expression is a factor indicating a poor prognosis for patients with NSCLC, and silencing of vimentin reduces the invasive ability of NSCLC cells. Vimentin can affect the prognosis of NSCLC by altering the invasive ability of cells. Vimentin in cancer cells might be a novel therapeutic target for NSCLC.
Footnotes
This article is freely accessible online.
- Received January 19, 2016.
- Revision received February 25, 2016.
- Accepted February 26, 2016.
- Copyright© 2016 International Institute of Anticancer Research (Dr. John G. Delinassios), All rights reserved





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}