


Abstract
Enhanced glycolysis in cancer cells presents a target for chemotherapy. Previous studies have indicated that proliferation of cancer cells can be inhibited by treatment with phenformin and with an inhibitor of 6-phosphofructo-2-kinase/fructose-2,6-bisphosphatase 3 (PFKFB) namely 3-(3-pyridinyl)-1-(4-pyridinyl)-2-propen-1-one (3PO). In the present work, the action of two inhibitors that are effective at lower concentrations than 3PO, namely 1-(3-pyridinyl)-3-(2-quinolinyl)-2-propen-1-one (PQP) and 1-(4-pyridinyl)-3-(2-quinolinyl)-2-propen-1-one (PFK15) were investigated. The inhibitors of lactate dehydrogenase (LDHA) studied in order of half-maximal inhibitory concentrations were methyl 1-hydroxy-6-phenyl-4-(trifluoromethyl)-1H-indole-2-carboxylate (NHI-2) < isosafrole < oxamate. In colonic and bladder cancer cells, additive growth inhibitory effects were seen with the LDHA inhibitors, of which NHI-2 was effective at the lowest concentrations. Growth inhibition was generally greater with PFK15 than with PQP. The increased acidification of the culture medium and glucose uptake caused by phenformin was blocked by combined treatment with PFKFB3 or LDHA inhibitors. The results suggest that combined treatment with phenformin and inhibitors of glycolysis can cause additive inhibition of cell proliferation and may mitigate lactic acidosis caused by phenformin when used as a single agent.
The tendency of cancer cells to exhibit increased rates of glycolysis was first noted by Otto Warburg in 1924 (1, 2). Enhanced glucose metabolism presents a target for therapeutic intervention if selective agents or combinations can be identified. The reaction catalyzed by phosphofructokinase is believed to be an important rate-limiting step in glycolysis and the enzyme is activated by fructose 2,6-bisphosphate. Concentrations of fructose 2,6-bisphosphate are regulated by bifunctional enzymes possessing phosphofructokinase 2 and fructose-2,6-bisphosphatase activities (PFKFBs). PFKFB3 is of particular interest due to its high rate of kinase to phosphatase activity and its increased level in cells of many cancer types (3,4). Studies in colonic cancer cells (5) and bladder cancer cells (6) indicate that a PFKFB3 inhibitor, 3-(3-pyridinyl)-1-(4-pyridinyl)-2-propen-1-one (3PO), can inhibit growth and can have additive effects with other agents that affect glucose metabolism. In the present work, two PFKFB3 inhibitors with effective concentrations lower than that of 3PO were examined as single agents and in combination with inhibitors of lactate dehydrogenase A (LDHA). LDHA was identified by several investigators as a potential target for therapeutic intervention in cancer cells (7, 8). Probably the most widely studied inhibitor of LDHA is oxamate although its half-maximal inhibitory concentration is high. More effective inhibitors of LDHA have been reported (9, 10) and in the present work were studied in combination with inhibitors of PFKFB3.
Biguanides exert a paradoxical effect on many cancer cells in that they appear to enhance glucose metabolism but can have growth-inhibitory effects. A synergistic anticancer effect of phenformin and oxamate has been reported (8). One of the goals of the present investigation was to extend previous studies with phenformin in order to characterize its combined action on cancer cell growth with compounds inhibiting glycolysis.
Materials and Methods
Reagents. Methyl 1-hydroxy-6-phenyl-4-(trifluoromethyl)-1H-indole-2-carboxylate (NHI-2) was purchased from Focus Biomolecules, Plymouth Meeting, PA, USA. 1-(3-pyridinyl)-3-(2-quinolinyl)-2-propen-1-one (PQP) was obtained from ChemBridge Corporation (San Diego, CA, USA). 1-(4-Pyridinyl)-3-(2-quinolinyl)-2-propen-1-one (PFK15), sodium oxamate, isosafrole and phenformin were obtained from Sigma-Aldrich (St. Louis, MO, USA).
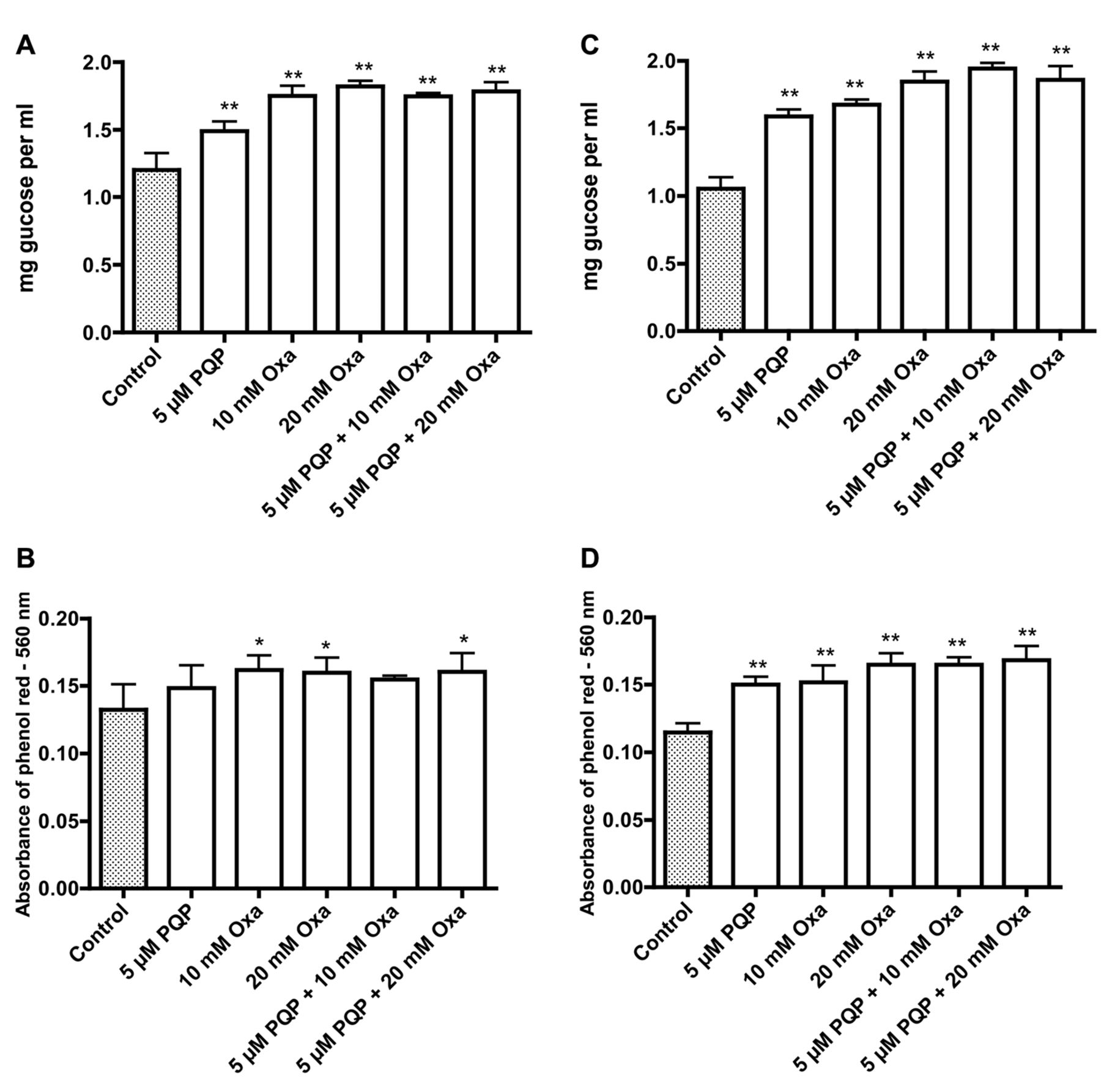
Effects of 1-(3-pyridinyl)-3-(2-quinolinyl)-2-propen-1-one (PQP) and oxamate (Oxa) on glucose concentration (A, C) in the medium and pH (B, D) in the medium of T24 (A, B) and UM-UC-3 (C, D) bladder cancer cells. Significantly different from the control at *p<0.05 and **p<0.01.
Cells and determination of growth. Human bladder cancer cell lines, namely 5637, HT1197, HT1376, RT4, SW780, T24, TCCSUP and UM-UC-3, and human colonic cancer cells (Caco-2, HCT116 and HT29) were obtained from the American Type Culture Collection (Rockville, MD, USA), and were incubated at 37°C with 5% carbon dioxide. After plating 5000 cells in 0.2 ml RPMI-1640 medium with 5% fetal calf serum for 24 hours, the medium was replaced with either control medium or medium containing drugs. Cell growth after a further 72 h was monitored by the increase in protein determined by staining with sulforhodamine B, essentially as described by Vichai and Kirtikara (11).
pH Determination. pH of the cell culture medium was monitored by measuring the light absorbance at 560 nm of phenol red in the medium. Decreasing absorbance is related to a lowering of the medium pH (12).
Glucose assay. Glucose was assayed with 2-μl cell culture medium using the glucose oxidase and peroxidase reagent from Sigma-Aldrich. This is a colorimetric procedure in which the oxidation of glucose is coupled with glucose oxidase and peroxidase to the oxidation of dianisidine. Incubation at room temperature was performed for 30 min in a total volume of 0.3 ml, including 80 μl 1 M sodium phosphate (pH 6.0). The incubation was stopped by the addition of 0.2 ml 12 N sulfuric acid. Light absorbance at 540 nm was measured with 0.2 ml of the mixture using a plate reader and the glucose concentration was calculated by reference to a glucose standard solution.
Statistical evaluation. Data are presented as means and standard deviations for six or a greater number of determinations. Statistical significance of the results was determined by Dunnett's test for multiple comparisons or by a two-tailed Student's t-test. A probability less than 5% was considered significant.

Effects of 1-(3-pyridinyl)-3-(2-quinolinyl)-2-propen-1-one (PQP) and oxamate (Oxa) on the growth of Caco-2 (A), HT29 (B), HT1197 (C), T24 (D) and UM-UC-3 (E) cancer cell lines. Significantly different from the control at **p<0.01.

Effects of 1-(3-pyridinyl)-3-(2-quinolinyl)-2-propen-1-one (PQP) and isosafrole (Iso) on glucose concentration (A, C) in the medium, pH (B, D) in the medium and growth of T24 (A-C) and UM-UC-3 (D-F) bladder cancer cells. Significantly different from the control at **p<0.01.

Effects of 1-(3-pyridinyl)-3-(2-quinolinyl)-2-propen-1-one (PQP) and 1-(4-pyridinyl)-3-(2-quinolinyl)-2-propen-1-one (PFK15) on the growth of Caco-2 (A), HT29 (B), RT4 (C) and UM-UC-3 (D) cancer cells. Significantly different from the control at *p<0.05 and **p<0.01.
Results
The data in Figure 1A and C show that the glucose concentration in the medium for T24 and UM-UC-3 bladder cancer cells decreased from an initial concentration of 2 mg per ml after a 72-h incubation. The uptake of glucose was diminished when the cells were incubated with PQP alone and sodium oxamate alone; the effect was greater with a combination treatment. Parallel changes in medium pH were indicated by the absorbance of phenol red at 560 nm (Figure 1B and D), and suggested that decreased uptake of glucose in the presence of PQP and oxamate is accompanied by decreased glycolysis and acidification of the medium. Results for the influence of PQP and oxamate on growth are presented for two colonic cancer cell lines (Caco-2 and HT29, Figure 2A and B) and three bladder cancer cell lines (HT117, T24 and UM-UC-3, Figure 2C-E). There was a tendency for the inhibition of growth to be greater with the combination treatment.
The results in Figure 3 illustrate the effects of treatment with PQP and an LDHA inhibitor that is effective at lower concentrations than oxamate, namely isosafrole (Figure 3). Concentrations of isosafrole that have been reported to inhibit lactate dehydrogenase (10) had only marginal effects on glucose uptake and medium pH for T24 and UM-UC-3 cells but for T24 cells there was a significant inhibition of growth with 0.5 mM isosafrole, and for both cell lines, the greatest inhibition of growth was seen with a combination treatment of PQP and 0.5 mM isosafrole (Figure 3C and F).
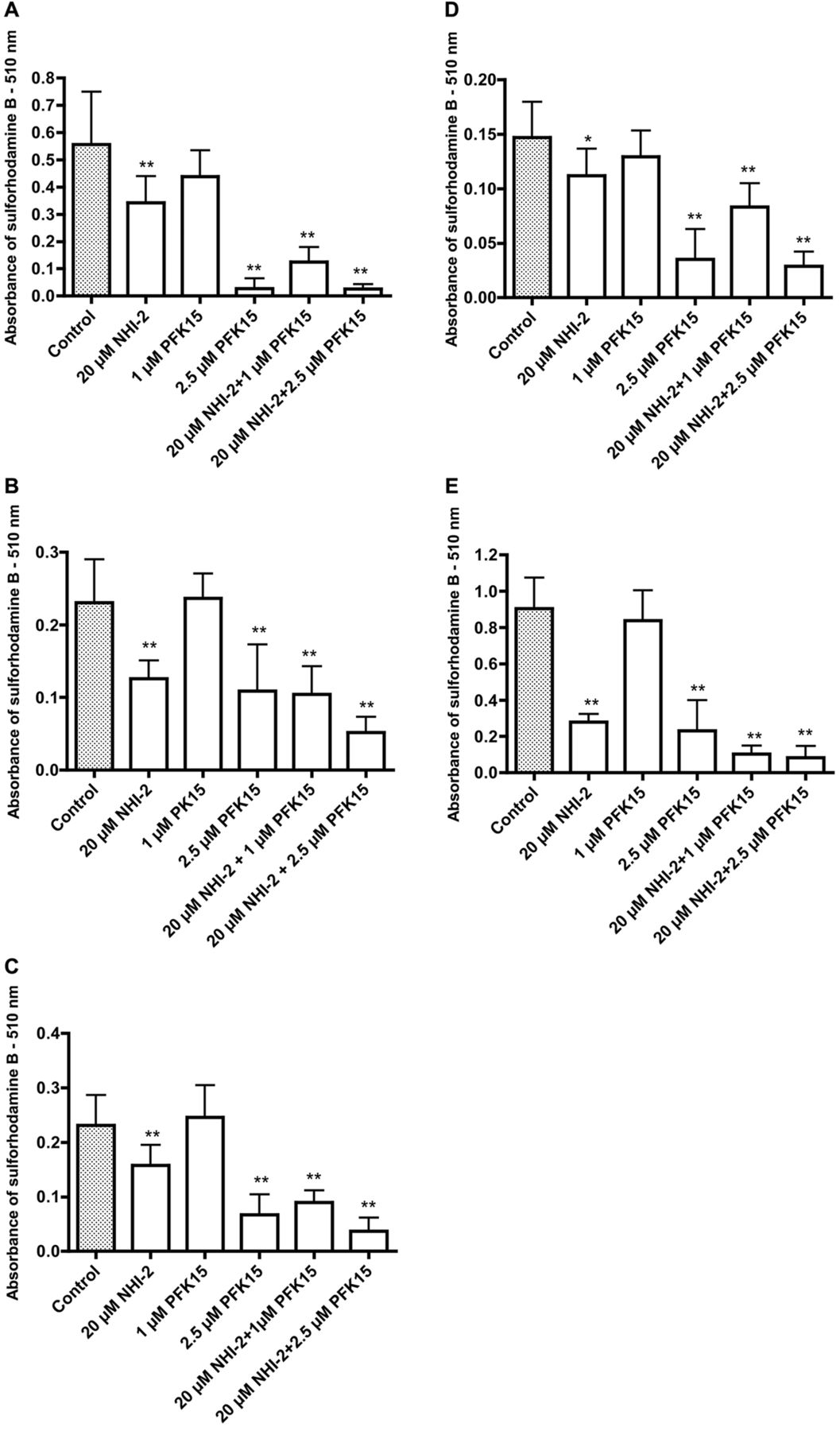
Effects of 1-(4-pyridinyl)-3-(2-quinolinyl)-2-propen-1-one (PFK15) and methyl 1-hydroxy-6-phenyl-4-(trifluoromethyl)-1H-indole-2-carboxylate (NHI-2) on the growth of HCT116 (A), HT1376 (B), SW780 (C) TCCSUP (D) and UM-UC-3 (E) cancer cells. Significantly different from the control at *p<0.05 and **p<0.01.

Effects of phenformin (Phen) and isosafrole (Iso) on glucose concentration (A, D) in the medium, pH (B, E) in the medium and growth (C, F) of HT29 (A-C) and T24 (D-F) cancer cells. Significantly different from the control at *p<0.05 and **p <0.01.

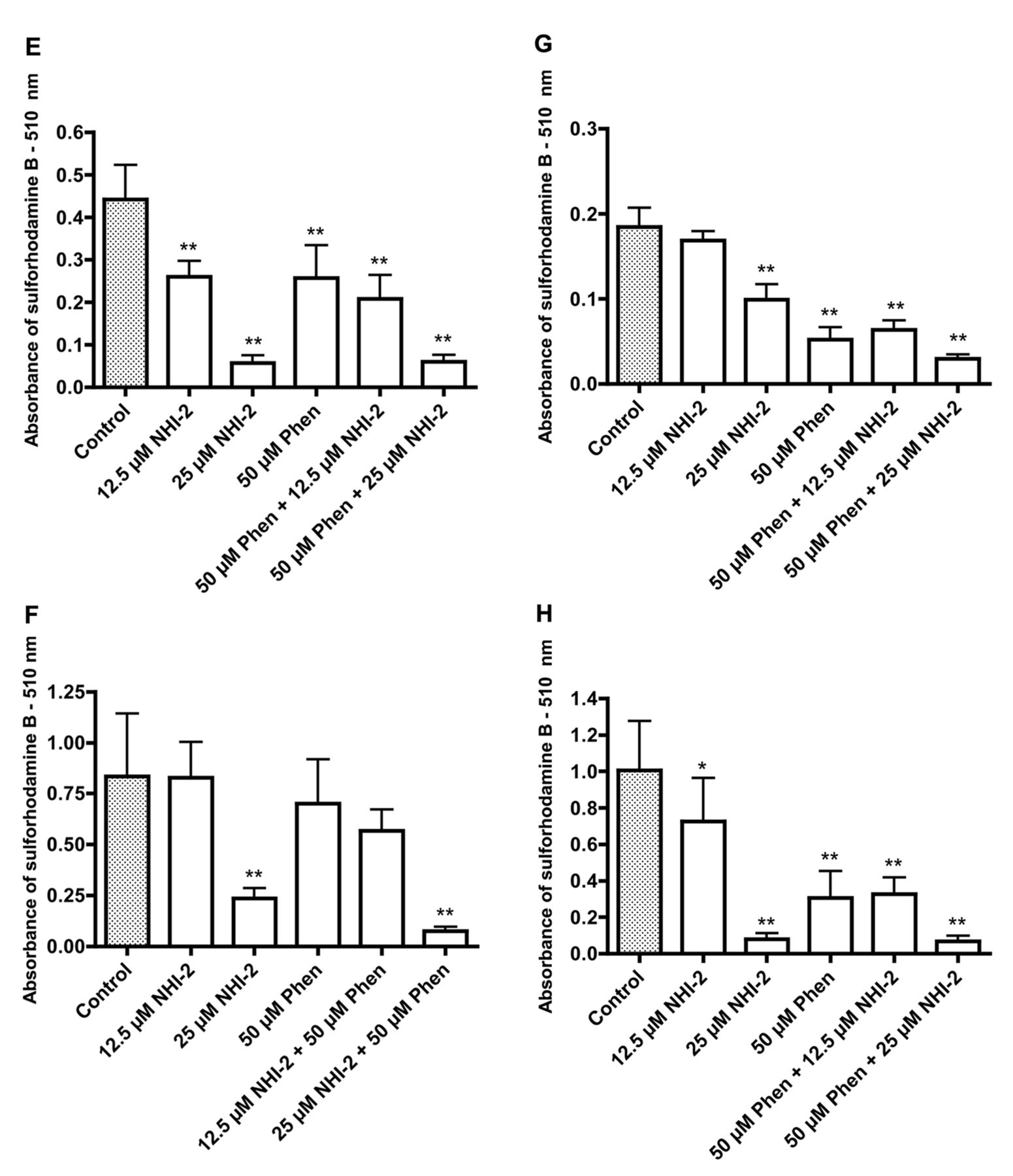
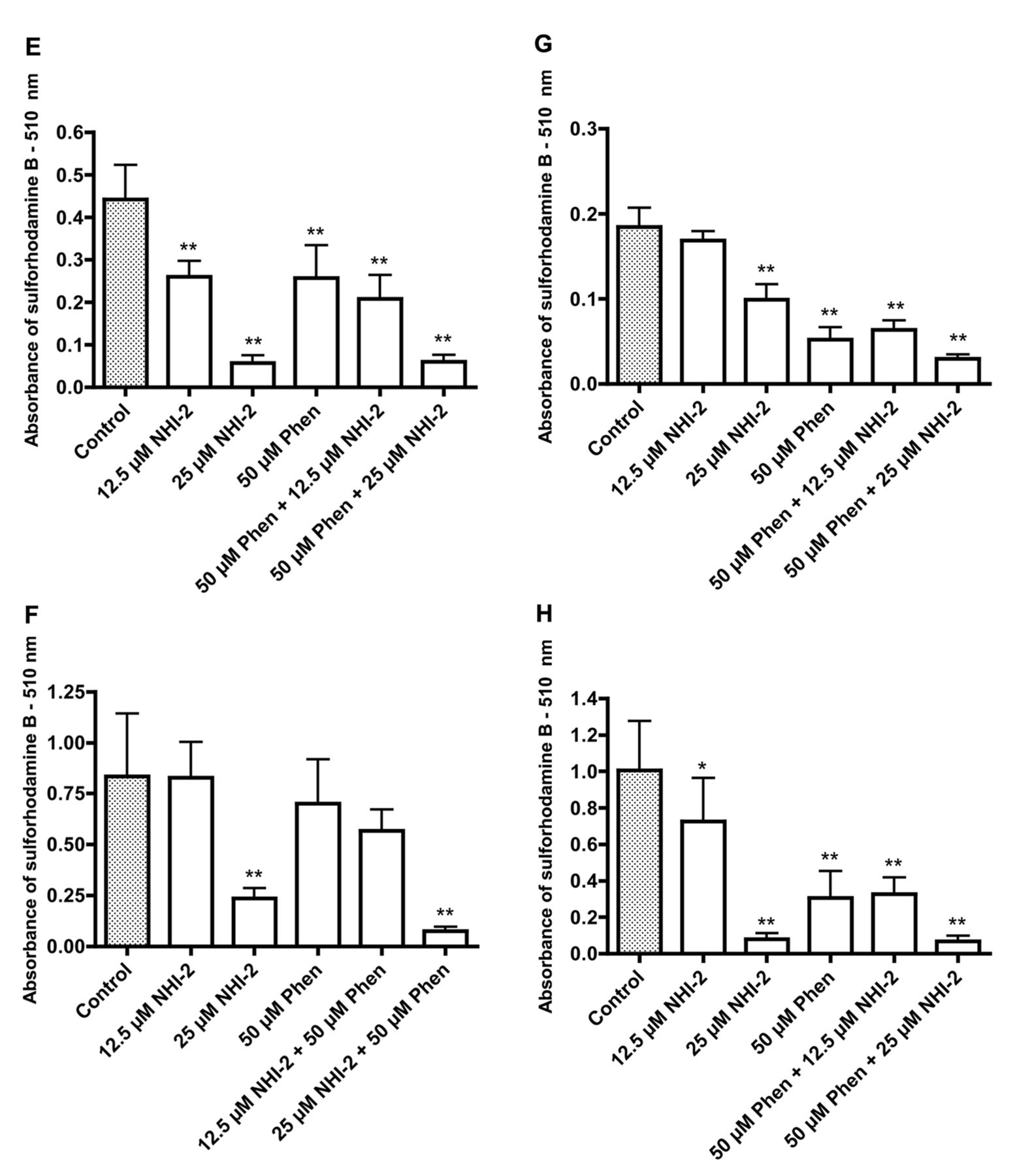
Effects of phenformin (Phen) and methyl 1-hydroxy-6-phenyl-4-(trifluoromethyl)-1H-indole-2-carboxylate (NHI-2) on the growth of the following cancer cell lines: 5637 (A), Caco-2 (B), HT29 (C), RT4 (D), SW780 (E), T24 (F), TCCSUP (G), and UM-UC-3 (H). Significantly different from the control at *p<0.05 and **p<0.01. Effects of phenformin (Phen) and methyl 1-hydroxy-6-phenyl-4-(trifluoromethyl)-1H-indole-2-carboxylate (NHI-2) on the growth of the following cancer cell lines: 5637 (A), Caco-2 (B), HT29 (C), RT4 (D), SW780 (E), T24 (F), TCCSUP (G), and UM-UC-3 (H). Significantly different from the control at *p<0.05 and **p<0.01.
The compounds PQP and PFK15 have a close structural relationship. The data in Figure 4 for two colonic cancer cells and two bladder cancer cell lines indicate that PQP and PFK15 have similar growth-inhibitory effects on these cell lines, with generally a somewhat greater effect with PFK15 after a 72-h incubation. The results in Figure 5 show the growth-inhibitory effects of PFK15 and one of the newer inhibitors of LDHA, namely NHI-2. Growth-inhibitory effects for both agents as single treatments are illustrated for a colonic cancer cell line (HCT116, Figure 5A) and four bladder cancer cell lines (HT1376, SW780, TCCSUP and UM-UC-3, Figure 5B-E). For all the cell lines, the greatest growth inhibition was seen with a combination treatment with NHI-2 and PFK15.
The data in Figure 6 show increased uptake of glucose after incubation of HT29 colon cancer cells and T24 bladder cancer cells with phenformin, whereas incubation with isosafrole was accompanied by decreased uptake of glucose that was paralleled by increased acidification of the medium with phenformin treatment and the converse with isosafrole. The results for the combination treatment suggested that treatment with isosafrole prevented the increased uptake of glucose and increased acidification of the medium caused by phenformin. Both phenformin and isosafrole caused growth inhibition with the possibility for some additive effects. The possibility of additive inhibitory effects on growth when cells are incubated with a combination of phenformin and an inhibitor of lactate dehydrogenase is further suggested by the data in Figure 7. Data for two colonic cancer cell lines and six bladder cancer cell lines indicate that the greatest growth inhibition was seen with a combination treatment with phenformin and NHI-2.
Discussion
Increased glycolysis is not a universal occurrence in cancer (13) but enhanced glucose uptake is observed in the majority of cases and is a useful diagnostic feature. High rates of glycolysis are most notable in rapidly growing tumors. Progress in directing chemotherapy against glycolytic enzymes has been restricted by the availability of selective drugs. A number of glycolytic enzymes have been seen as potential targets, including hexokinase, phosphofructokinase, glyceraldehyde-3-phosphate dehydrogenase, pyruvate kinase and lactate dehydrogenase (9, 14, 15). The activity of phosphofructokinase is stimulated by fructose 2,6-bisphosphate. This has encouraged the search for inhibitors of PFKFBs, the enzymes that determine the levels of fructose 2,6-bisphosphate. PFKFB3 has been investigated on the basis that the ratio of kinase to phosphatase activity is high and the enzyme is frequently up-regulated in cancer (3).
Combination chemotherapy against cancer is a widespread approach, with the rationale of limiting the toxicity of single agents and rendering resistance more difficult for the cancer cell. In the present work, combinations of inhibitors of PFKFB3 and LDHA were investigated. Structure–activity studies have suggested that the substitution of a quinolinyl group for a pyridine ring in 3PO can yield more effective inhibitors of PFKFB3 (3). Two such compounds were studied in the present work, with PFK15 proving somewhat more growth-inhibitory than PQP. Initial studies were performed with oxamate which is a widely studied LDHA inhibitor. Experiments with more recently identified LDHA inhibitors, isosafrole and NHI-2, indicated their greater potency as growth inhibitors of bladder and colon cancer cells. Additive, although not dramatically synergistic, effects were observed with combinations of LDHA and PFKFB3 inhibitors.
Inhibition of glycolysis in cancer cells may force them to become more dependent on oxidative phosphorylation for ATP production. This provides a rationale for combining treatment with inhibitors of glycolysis and oxidative phosphorylation. The work of Ben Sahra et al. using a combination of metformin and 2-deoxyglucose may be considered an example of this approach (16). Metformin is a weaker inhibitor of oxidative phosphorylation and growth than the structurally related biguanide, phenformin. Combination treatment with phenformin and oxamate was reported by Miskimins et al. (8). The present work suggests that more effective LDHA inhibitors, notably NHI-2, may be advantageous in such an approach. Although still used in some countries, phenformin was withdrawn in the 1970s in the United States for the therapy of type 2 diabetes after reports of cases of fatal lactic acidosis. The data suggesting that combination treatment with inhibitors of glycolysis have the potential to mitigate this action of phenformin may encourage further examination of phenformin as a therapeutic agent in cancer therapy.
Acknowledgements
This work was supported by the Alma Toorock Memorial for Cancer Research.
Footnotes
This article is freely accessible online.
- Received January 19, 2016.
- Revision received February 27, 2016.
- Accepted March 4, 2016.
- Copyright© 2016 International Institute of Anticancer Research (Dr. John G. Delinassios), All rights reserved
References
In this issue





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Jump to section
Related Articles
Cited By...
- Galloflavin Plus Metformin Treatment Impairs Pancreatic Cancer Cells
- Effects of Biguanides on Growth and Glycolysis of Bladder and Colon Cancer Cells
- A New Prognostic Tool for Patients Undergoing Radiotherapy plus Upfront Transurethral Resection for Bladder Cancer
- Simplified Comorbidity Score and Eastern Cooperative Oncology Group Performance Score Predicts Survival in Patients Receiving Organ-preserving Treatment for Bladder Cancer
- A New Prognostic Instrument Specifically Designed for Patients Irradiated for Recurrent Carcinoma of the Bladder
- A Survival Score for Patients Assigned to Palliative Radiotherapy for Metastatic Bladder Cancer
- Concurrent Chemotherapy Improves the Overall Survival of Patients Irradiated for Locally Recurrent Bladder Cancer