


Abstract
Background/Aim: The aim of this study was to evaluate the expression of FASL, FAS and FADD and caspase-3 in oesophagus, stomach and colonic tissues of mice irradiated in vivo by immunohistochemistry. Materials and Methods: A total of 48 adult male C57BL mice were distributed into four groups: Ami−/Rad−: Mice received 0.5 ml of 0.9% physiological saline solution (PPS) intraperitioneally (i.p.); Ami+/Rad−: mice received amifostine (400mg/kg i.p.) freshly dissolved in double-distilled water; Ami−/Rad+: mice received 0.5 ml of PSS i.p. 30 min before a single whole-body radiation dose of 7 Gy; Ami+/Rad+: mice received 0.5 ml of an aqueous solution of 400 mg/kg amifostine i.p.30 min prior to irradiation. All groups were assigned into subgroups sacrificed at 0.5 h, 1 h, 2 h and 4 h after irradiation. Results. In oesophagus and stomach tissues, we did not observe any difference between Ami−/ad−, Ami+/Rad−, Ami−/Rad+ and Ami+/Rad+ groups in the expression of FASL, FAS and FADD. The colonic tissue was the only to exhibit any difference in the expression of FAS and caspase-3 protein in the Ami−/Rad+group at 1 and 2 h. Amifostine increased FAS and caspase-3 immunoexpression when compared to the control. Immunoexpression for FASL and FADD was not remarkably different in colonic tissue. Conclusion: Taken together, our results demonstrate that amifostine increases FAS and caspase-3 expression in colonic tissue of irradiated mice.
Radiation toxicity is an important problem that limits treatment intensity and adversely affects patients' quality of life. In this context, the study of drugs for manipulating the cellular response to radiation is of great interest. The compound WR-2721 was developed by the U.S. Army Anti-Radiation Drug Development Program for its potential to protect against damage caused by ionizing radiation (1). Today, WR-2721 is known as amifostine (Ethyol;Med Immune Oncology, Inc., Gaithersburg, MD,USA) and its use was approved by the Food and drug Administration in 1995 to reduce specific chemotherapy-induced toxicities. When given at high doses before exposure to radiation, WR-2721 protected mice against radiation-induced tumors (2, 3). On the other hand, ionizing radiation has been shown to induce apoptotic cell death of bone marrow cells shortly after radiation exposure (4). Amifostine has been shown to protect preferentially normal cells from caspase-3 activation and apoptosis after anticancer drugs and irradiation in contrast to tumor cells (5, 6).
Apoptosis or programmed cell death is an efficient physiological mechanism for cell suicide that is important for growth and cell differentiation processes in multi-cellular animals, controlling the number of cells and organ size (7). Execution of apoptosis is a complex, tightly regulated and active cellular process whereby individual cells are triggered to undergo self-destruction in a manner which will neither injure neighboring cells nor elicit aninflammatory reaction (8). One of the characteristics of the great majority of cancers is their progressive resistance to apoptosis.
The morphological alterations observed in apoptosis are induced by the action of the caspases, a family of cysteine proteases that constitute the central regulatory mechanism of cell death by apoptosis. Caspase-3 is the principal trigger of the apoptotic process. Extrinsic and intrinsic pathways promote the activation of the caspase cascade.
The extrinsic apoptotic pathway following FAS (also known as APO1 or CD95) binding has been well-characterized (9, 10). FAS ligand (FASL) is a type II membrane protein. FAS is a 48-kDa type I membrane protein, with a cysteine-rich extracellular domain of 155 amino acids, which is expressed in various rapidly proliferating cells, including thymocytes, immunocompetent T-lymphocytes, and epithelial cells of the skin and gut (11). Oligomerization of FAS by FASL recruits the adaptor molecule FAS-associated death domain (FADD) to the death domain of the FAS intracellular region (12). Procaspase-8, which is an inactive zymogen, associates in turn with FADD by interaction between their death effector domains (13). Within the FAS-FADD-procaspase-8 complex, called the death-inducing signaling complex (DISC) (14), procaspase-8 undergoes auto-cleavage to be converted into an active form which activates downstream effector caspase-3 (11). FAS-mediated apoptosis is involved in several inflammatory diseases, autoimmune diseases and cancer (15-17).
Mean scores of FASL immunoexpression in the digestive tract of mice treated with amifostine with/without irradiation.
Several mechanisms have been suggested for the radioprotective effects of amifostine, including free radical-scavenging, auto-oxidation leading to intracellular hypoxia, chemical repair by hydrogen atom donation and also due to its ability to modulate the transcriptional regulation of genes that are involved in apoptosis, cell cycle and DNA repair (18, 19).
Continuing another study of our research group (20), we analyzed the expression of FASL, FAS and FADD by immunohistochemistry in the oesophagus, stomach and colon of mice irradiated in vivo.
Material and Methods
Animals. A total of 48 adult male C57BL mice (20-24g) were used. The study was approved by the Ethical Committee, Federal University of São Paulo (Sao Paulo, SP, Brazil, number 0134/2011), and the animals used received humane care (20). During the experiments, the mice were kept in plastic cages on sawdust, with rations and tap water available ad libitum. Animals were randomly assigned to the following groups: Ami−/Rad−: Twelve non-irradiated mice received 0.5 ml of 0.9% physiological saline solution (PPS) intraperitioneally (i.p.). They were distributed into four sub-groups and sacrificed by cervical dislocation at 0.5 h (n=3), 1 h (n=3), 2 h (n=3) and 4 h (n=5). Ami+/Rad−: Twelve non-irradiated mice received amifostine (400mg/kg i.p.) freshly dissolved in double-distilled water. Mice were assigned into subgroups and sacrificed at the same time intervals as in the first group. Ami−/Rad+: Twelve mice received 0.5ml of PSS i.p.30min before a single whole-body radiation dose of 7 Gy. They were assigned to four subgroups and sacrificed at 0.5, 1, 2 and 4 h after irradiation.
Mean scores of FAS immunoexpression in the digestive tract of mice treated with amifostine with/without irradiation.
Ami+/Rad+: Twelve mice received 0.5 ml of an aqueous solution of 400 mg/kg amifostine i.p. 30 min prior to irradiation and were assigned into subgroups and sacrificed at the same time intervals as in the previous groups.
Radiation factors. A60Co radiation source (Teletherapy machine - Alcion II, CGR, Paris, France) with a dose rate of 1.69 Gy min−1 was used. The mice received a single whole-body radiation dose of 7Gy. During irradiation, the animals were placed in a 20×20cm wooden box with a plastic cover and internal divisions to allow immobilization, without anesthesia.
Immunohistochemistry. Conventional 3 μm-thick sections were obtained and mounted on slides pretreated with 3-minopropyltriethoxysilane (Sigma St. Louis, USA). Sections were then deparaffinized, hydrated and processed as follows. For antigen retrieval slides were placed in 0.01 M citrate-buffer (pH 6.0) and heated in a steamer for 30 min. Endogenous peroxidase was blocked by using 10% hydrogen peroxide for 20 min. After overnight incubation of sections with rabbit polyclonal antibody to FASL(C-178, 1:100; Santa Cruz, San Diego, California USA); rabbit polyclonal anti-FAS(FL-335, 1:100; Santa Cruz), goat polyclonal anti-FADD (S-18, 1:150; Santa Cruz) and polyclonal cleaved caspase-3 (Asp 175) (1:300;Cell Signaling, NY, USA). Sections were allowed to react with secondary biotinylating antibody and streptavidin-biotin-peroxidase (LSAB; Dakocytomation, Copenhagem, Denmark) for 30 min each. Finally, the immunostaining reaction was revealed using 3,3’-diaminobenzidinetetrahydrocloride (Sigma) (0.07 diaminobenzidine in PBS buffer 0.05 M, pH 7.6, 0.01% hydrogen peroxide) counterstained with Harris's hematoxylin and coverslipped with Entellan (Sigma). Negative and positive controls were made to run simultaneously. Negative controls were made by eliminating the primary antibody; positive control was represented by gastric adenocarcinoma tissue.
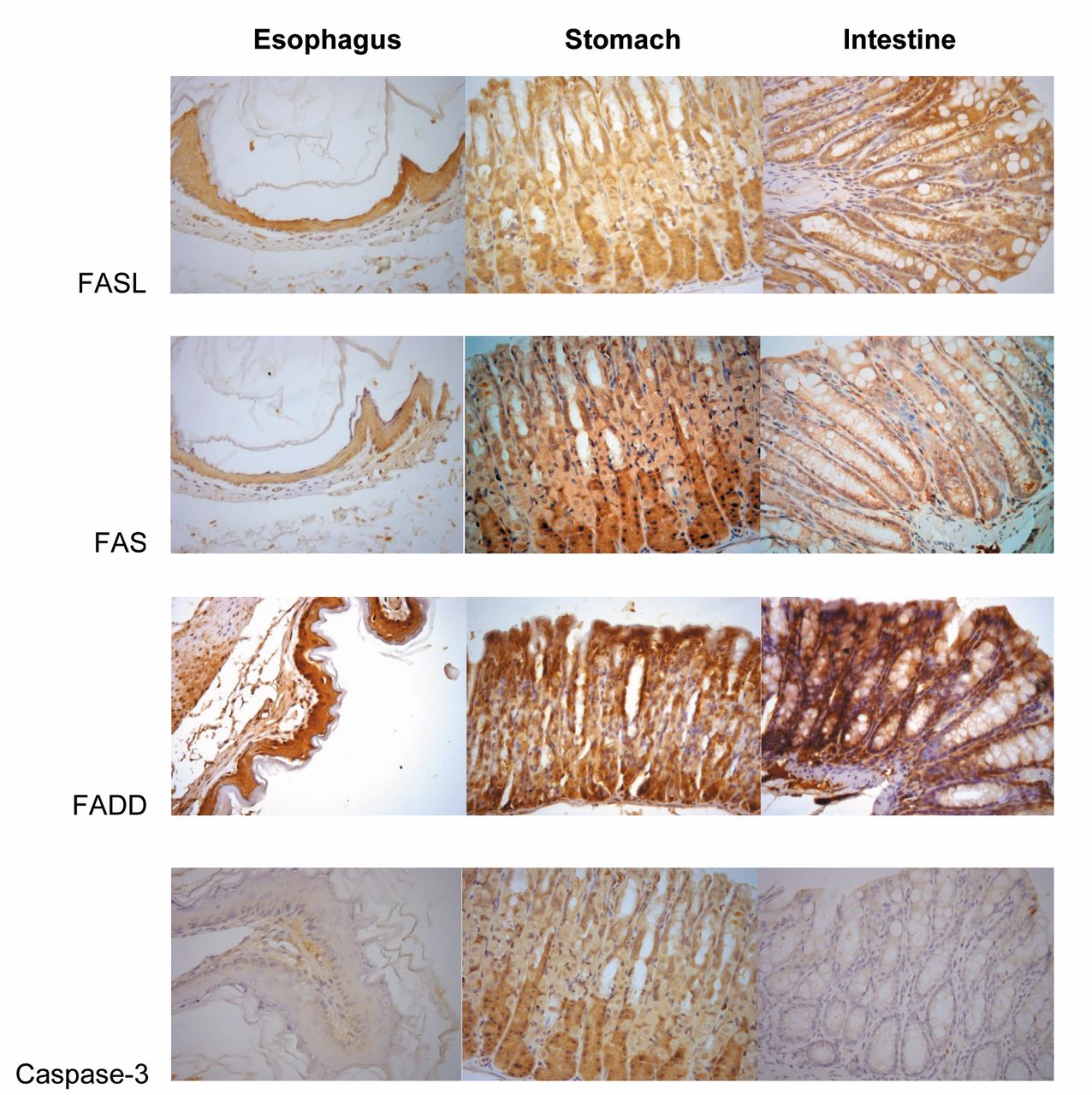
FASL, FAS, FADD and cleaved caspase-3 expression assessment using the immunohistochemical streptavidin-biotin-peroxidase method. Oesophagus, stomach and colon showed strong cytoplasmic staining patterns (score 4). Cleaved caspase-3 was not detected or was faintly positive. The immunohistochemical assessment was performed at a magnification of ×400.
Evaluation criteria.The pattern of positivity for FASL, FAS and FADD was predominantly the cytoplasmic type. This pattern was analyzed according to the criteria of distribution and intensity. A numerical scoring system with two categories was used to assess the observed expression of the proteins. Category A documented the number of immunoreactive cells as 0 or negative (no immunoreactive cells or <10% immunoreactive), 1 (≥10% to <50% immunoreactive), or 2 (≥50% immunoreactive). Category B documented the intensity of the immunostaining as 1 (no immunostaining or weak), or 2 (moderate or strong). The values for categories A and B were added to give the immunoreactivy score, which could range from 0 to 4. Zero was considered negative, 1 was considered weak, 2 moderate and 4 strong staining. Representative areas were photographed by digital camera under ×400 magnification.
Mean scores of FADD immunoexpression in the digestive tract of mice treated with amifostine with/without irradiation.
Statistical methods.The results were submitted to statistical analysis using the Kruskall-Wallis test followed by Dunn's test. p-Values of less than 0.05 were considered statistically significant.
Results
The pattern of positivity for FASL, FAS and FADD was predominantly of cytoplasmic type with strong intensity (Figure 1). In oesophageal and stomach tissues, we did not observe any difference between Ami−/Rad−, Ami+/Rad−, Ami−/Rad+ and Ami+/Rad+ groups for the expression of FASL, FAS and FADD. No significant association was observed between FASL, FAS and FADD expressions at the studied periods (Tables I and II).
The colonic tissue was the only one that showed differences in the expression of FAS protein in the Ami−/Rad+ group at 1, 2 and 4 h. At 0.5 h, we did not observe any difference in the expression of FAS. Amifostine increased FAS immunoexpression when compared to the control. As in the esophageal and stomach tissue, we did not observe differences between Ami−/Rad−, Ami+/Rad−, Ami−/Rad+ and Ami+/Rad+ groups in the expression of FASL and FADD (Table III).
The expression of cleaved caspase-3 did not differ in the stomach and esophagus for any of the periods of study. Nevertheless, amifosfine increased caspase-3 expression at 0.5 h and 1 h after radiation exposure. Such findings are summarized in Table IV.
Mean scores of cleaved caspase-3 immunoexpression in the digestive tract of mice treated with amifostine with/without irradiation.
Discussion
The main damage induced by ionizing radiation concerns DNA, however, cytoplasmic organelles are also damaged by radiation (21). Ionizing radiation at dose levels used in clinical radiotherapy leads to cell death through induction of apoptosis. In this sense, it is very important to investigate the effects of whole-body radiation and the protective effect of amifostine on different body systems.
Amifostine is a pro-drug (WR-2721) converted by alkaline phosphatase to an active de-phosphorylated form (WR-1065) which penetrates the cell membrane by both active and passive diffusion. Amifostine accumulates more rapidly in normal tissues, which has a greater ability to dephosphorylate the drug to the free thiol due to the more neutral intracellular environment than that of some tumors (22). Consequently, it preferentially protects normal cells as opposed to tumors. For this reason, there is a strong clinical interest in the role of amifostine in the cell signaling pathways.
In the current study, none of the three tissues analyzed were different in FasL expression. There were no differences between the study periods. However, the colonic tissue showed differences in the expression of FAS at 1, 2 and 4 h. Amifostine increased FAS expression in colonic tissue of irradiated mice; in half an hour there was no difference. Esophagus and stomach also revealed no differences with respect to this protein. FADD protein expression did not differ in the three tissues.
FAS and FASL have been identified as very important effectors of apoptosis in various biological conditions, and the roles of these proteins are potentially important for elucidating the response of tumors to ionizing radiation.
Sbih-Lammali et al. have shown that nasopharyngeal carcinoma cells (NPC) constitutively express high levels of FAS, which retains its ability to transduce a potent signal of apoptosis (23). On the other hand, Abdulkarin et al., examining a typical EBV-positive xenografted NPC tumor line found that FASL expression, which was almost undetectable in NPC cells in the absence of irradiation, was dramatically increased after radiation exposure (24). The authors suggested that induction of FasL can mediate radiation-induced apoptosis in NPC.
Our results are in agreement that the tissues of the esophagus and stomach maintain functional FAS, fully capable of signal transduction of apoptosis. On the other hand, decreasing FAS expression in the intestine 1 h after irradiation may be due to apoptosis signaling process.
Transfected cell line MCF-7-FAS presents ultraviolet radiation-induced apoptosis mediated by activation of FAS/APO1 in a time- and dose-dependent manner (25). Despite the different type of radiation, in our study whole-body irradiation produced differences in the expression of FAS protein in colonic tissue. The same was true for treatment with amifostine, in which the xenobiotic increased FAS expression. Therefore, it was possible to evaluate the role of amifostine on FAS protein in the oesophagus, stomach and colon and at different times of study. Oligomerization of FAS by FASL recruits the adaptor molecule FADD. UV-induced apoptosis is protected by the dominant negative form of FADD (25). In the same way, whole-body irradiation did not provoke any difference in the expression of FADD protein. The same was true for treatment with amifostine.
Recently, Ormsby et al. investigated whether amifostine stimulates apoptosis when administered after radiation exposure (26). Mice were administered 400 mg/kg of amifostine 30 min before or 3 h after whole-body irradiation with 0.25 (the maximum radiation dose for radiation emergency workers) or 6 Gy X-rays (high dose relevant to radiotherapy exposures) and apoptosis was analyzed 3 or 7 h later in spleen and bone marrow. A significant increase in radiation-induced apoptosis in the spleen of mice was observed when amifostine was administered before or after 0.25 Gy X-rays. In contrast, when a high dose of radiation was used (6 Gy), amifostine caused a reduction in radiation-induced apoptosis 3 h post-irradiation in spleen and bone marrow. This shows that amifostine can reduce apoptosis due to high doses of radiation but it does not mediate the same effect in response to low-dose exposures.
The immunostaining of cleaved caspase-3 was mostly undetected in the cytoplasm of all tissues studiedhere. Our results demonstrate that amifosfine was able to trigger apoptosis as a result of increasing caspase-3 immunoexpression in colonic cells 30 min and 1 h after radiation. Amifostine has been shown to protect preferentially normal cells from caspase-3 activation and apoptosis after anticancer drugs and irradiation as opposed to tumor cells (6,20). Amifostine reduces radiation damage in bone marrow cells, but does not completely inhibit it (20).
The results of FASL, FAS and FADD expression in the oesophagus, stomach and colon indicate that apoptosis is activated by the extrinsic pathway. At this stage, there was probably inhibition of apoptosis According to the literature, the presence of c-FLIP (FADD-like IL-1β-converting enzyme)-inhibitory protein) can inhibit the activation of caspase-8 by containing the effector domain of death, competing with caspase-8 for binding to FADD. Therefore, the lack of detection of expression of cleaved caspase-3 suggests the decrease of apoptosis signalling through caspase-8 activity.
Footnotes
-
Conflicts of Interest
None declared.
- Received January 20, 2015.
- Revision received February 6, 2015.
- Accepted February 10, 2015.
- Copyright© 2015 International Institute of Anticancer Research (Dr. John G. Delinassios), All rights reserved





{kind=link}