


Abstract
Background: Oil palm phenolics (OPP) or Palm Juice (PJ), a water soluble extract from the palm fruit (Elaies guineensis) has been documented to have anti-carcinogenic activities in various cancer types. Materials and Methods: To investigate OPP effects in pancreatic cancer (PaCa) cells, two PaCa cell lines (PANC-1 and BxPC-3) were treated with different OPP doses. The anti-proliferative, apoptotic and anti-invasive properties of OPP were evaluated using MTS, cytoplasmic histone-DNA fragmentation and matrigel invasive assays, respectively. Results: OPP suppressed PaCa proliferation in a dose-dependent manner. Its anti-invasive effects were validated by decreased expressions of MMP-9 and VEGF. Cell-cycle analysis demonstrated that cells were arrested in the S phase. OPP-induced apoptosis was associated with decrease in survivin and Bcl-XL expressions and increased expression of cleaved caspase-3, caspase-9 and PARP. Conclusion: Overall, our results demonstrate the anti-tumor effects of OPP on PaCa cells, providing initial evidence towards its potential therapeutic use.
Pancreatic cancer (PC) is the fourth leading cause of cancer deaths in the United States with a median survival of less than six months. Approximately 40,000 individuals were diagnosed with pancreatic cancer in 2010 and the number of estimated new cases has been increasing every year (1). Despite more focused research in this field in recent years, PC continues to be a therapeutic challenge. The incidence and mortality rates of this lethal disease have not yet shown a declining trend (2). Currently, the only known cure for PC diagnosed in the early stages is pancreatectomy (3). Other treatment modalities, such as chemo-, targeted- and radiotherapies may also be utilized in later stages of the disease. With regard to chemotherapy, current chemotherapeutic agents include gemcitabine (a purine analog), fluoropyrimidine and/or platinum drugs. (3). However, the major obstacle for these anticancer drugs is high toxicity with poor clinical results. Erlotinib, an epidermal growth factor receptor (EGFR) tyrosine kinase inhibitor, which is used as an adjuvant to gemcitabine, only shows marginally-improved survival benefits (4).
Due to poor outcomes with conventional treatments, novel strategies and drugs are needed for more effective treatment. One treatment strategy extensively investigated in pancreatic cancer research is the targeting of the nuclear factor-κB (NF-κB) signaling pathway. NF-κB is a ubiquitously expressed transcription factor that is responsible for mounting inflammatory responses due to stress. Moreover, NF-κB can be activated through a wide variety of stimuli such as, bacteria, viruses, UV radiation, phenobarbital, mitogens, growth factors, hormones, physical and oxidative stress (5). When stimulated, NF-κB is translocated to the nucleus where it can bind to promoters of its target genes to begin transcription of specific genes (6). NF-κB has been shown to be constitutively activated in approximately 67% of pancreatic adenocarcinomas compared to the normal pancreas tissue. This is associated with aggressive stage pancreatic cancer and drug resistance (7). Additionally, NF-κB downstream gene over-expressions are involved in apoptosis resistance, angiogenesis, enhanced invasion and metastasis, which intensify the aggressiveness of this disease and further complicate its treatment (8). As of now, however, agents that directly target NF-κB are not available.
It is widely known that consumption of diets containing high amounts of phytochemicals can offer protection against many types of cancers due to their antioxidant properties. Many epidemiological studies have proven this by consistently showing that regular consumption of plant foods, which are rich in phytochemicals, markedly reduce the risk of cancer (9). From many types of antioxidants that are naturally occurring in plants, the two most important that have been identified with many therapeutic benefits include carotenoids and phenolics (10).
The fruit of the oil palm, the major global palm oil crop, is chiefly used for extraction of edible oils and accounts for 33% of vegetable oil and 45% of edible oil worldwide (11). In addition to the oil, the palm fruit (Elaies guineensis), contains large quantities of phytochemicals such as, vitamin E (tocopherols, tocotrienols), carotenoids and phytosterols, which can be lipid- or water-soluble (12). The lipid-soluble compounds found in oil palm, especially tocotrienols, have been shown to have anti-tumor effects on different human cancer cells including prostate, breast, colon, melanoma and lung cancers (13-15). Recently, the water-soluble components of oil palm, called palm juice (PJ), which consist mainly of phenolic acids, demonstrated potential nutraceutical properties. Thus Palm juice is also referred to as oil palm phenolics (OPP). Crude water and ethanol palm fruit extracts have been shown to be able to scavenge free radicals by either hydrogen or electron donating mechanisms and can, therefore, act as primary antioxidants (16). More recent in vitro findings have revealed that OPP can inhibit proliferation and induce apoptosis in different cancer cell lines, such as breast, lung and skin cancers (15). Similarly, in animal models, it was also found that OPP has the ability to reduce tumor growth (17, 18). Given the critical role of NF-κB in modulating carcinogenic activity in pancreatic cancer combined with available data from past research on OPP, in this work, we investigated the potential of OPP as an anti-carcinogenic agent in pancreatic cancer cells via down-regulation of the NF-κB pathway.
Materials and Methods
Cell culture, reagents and antibodies. The human PC cell lines, PANC-1 and BxPC-3, obtained from ATCC (Manassas, VA, USA) were grown in DMEM medium (Mediatech, Manassas, VA, USA) supplemented with 10% fetal bovine serum and 1% penicillin and streptomycin in 5% CO2. OPP was provided by the Malaysian Palm Oil Board (Malaysian Palm Oil Board, Kuala Lumpur, Malaysia) at a stock concentration of 1,500 mg/ml gallic acid equivalents (GE). The protease inhibitor cocktail was obtained from Sigma (St. Louis, MO, USA). Primary antibodies for Poly (ADP-ribose) polymerase (PARP), β-actin and cell lysis buffer (20 mM Tris-HCl pH 7.5, 150 mM NaCl, 1 mM Na2EDTA, 1 mM EGTA, 1% Triton, 2.5 mM sodium pyrophosphate, 1 mM beta-glycerophosphate, 1 mM Na3VO4, 1 μg/ml leupeptin) were purchased from Cell Signaling Technology (Danvers, MA, USA). Primary antibodies against cleaved caspase-3 and cleaved caspase9, survivin, Bcl-XL were bought from Santa Cruz Biotechnology (Santa Cruz, CA, USA). The secondary antibodies were bought from Bio-Rad Laboratories (Hercules, CA, USA).
Cell viability studies by the MTS assay. The PANC-1 and BxPC-3 cells (5×103) were seeded in 96-well culture plates. After overnight incubation, the medium was removed and replaced with a fresh medium containing DMSO (vehicle control) or different concentrations of OPP. After 72 h of incubation, 20μl of CellTiter 96 AQueous One Solution Reagent (Promega, Madison, WI) was added to each well. After a 2-h incubation at 37°C in a humidified, 5% CO2 atmosphere, the absorbance at 490 nm was recorded on ELx800 plate reader (Bio-Tek; Winooski, VT, USA). Each variant of the experiment was performed in triplicate.
Histone/DNA ELISA for detection of apoptosis. The Cell Death Detection ELISA Kit (Roche, Palo Alto, CA, USA) was used to detect apoptosis in PC cells as previously described (19). Briefly, 105 cells were seeded in six-well plates. After 24 h of incubation, cells were treated with OPP or control for 72 h. The cells were then lysed and cytoplasmic histone/DNA fragments were extracted and incubated in microtiter plate modules coated with anti-histone antibody. In order to detect the immobilized histone/DNA fragment, peroxidase-conjugated anti-DNA antibody was used before color development with ABTS substrate for peroxidase. The spectrophotometric absorbance of the samples was determined by using an ELx800 plate reader (Bio-Tek) at 405 nm.
Clonogenic assay. One million cells were seeded in a 100-mm dish and incubated overnight. Subsequently, the cells were cultured with OPP or control and grown for 72 hours. Subsequently, viable cells were counted and plated in 100 mm dishes in a range of 5,000 cells per plate. The cells were then incubated for 21 days at 37°C in a 5% CO2 incubator. All the colonies were fixed in 4% paraformaldehyde and stained with 2% crystal violet.
Flow cytometry and cell-cycle analysis. Cells were seeded in a 100 mm dish and incubated overnight. Subsequently, all the cells were starved for another 24 h. The cells were released into control or OPP containing media and grown for 72 hours. Then, the cells were collected and fixed with ice-cold 70% (v/v) ethanol for 24 h. After centrifugation at 3000 ×g for 5 min, the cell pellets were washed with PBS (pH 7.4) and re-suspended in PBS containing propidium iodide (PI; 50 μg/ml) and DNase-free RNase (1 μg/ml). Samples were then incubated at room temperature for 2 h and the DNA content was determined by flow cytometry using a FACScan flow cytometer (BD; San Jose, CA, USA).
Annexin V-FITC method for apoptosis analysis. The Annexin V-FITC apoptosis detection kit (BD) was used to measure the apoptotic cells. Briefly, PANC-1 and BxPC-3 cells were incubated in the presence or absence of OPP for 48 h. Cells were trypsinized, washed twice with ice-cold PBS and re-suspended in 1 × binding buffer at a concentration of 105/ml cells in a total volume of 100 μl. After that, 5 μl of Annexin V-FITC and 5 μl of PI were added. All the samples were kept in the dark for 20 min at room temperature. Finally, 400 μl of 1 × binding buffer was then added to each tube and the number of apoptotic cells was analyzed by flow cytometry (BD).
Wound healing assay. PANC-1 and BxPC-3 were seeded in a six-well plate at a concentration of 4×105 cells per well. After overnight incubation, the culture media were removed and a scratch wound across each well was made using fine tips. All the wound areas were washed three times with PBS to make sure no loosely held cells were attached. Subsequently, the cells were cultured in the presence or absence of OPP and the wound images were taken as 0 hour. After 20 hours, wound healing pictures were taken under a microscope.
Cell invasive assay. The BD Biocoat invasion kit (BD) was used to evaluate the tumor invasive ability according to the manufacturer's protocol. Briefly, around 5×104 cells of PANC-1 and BxPC-3 with basal media were transferred in each 6-well upper chamber in the presence or absence of OPP. In the meantime, 3 mm of culture medium with 10% FBS was added into each lower chamber of a 24-well plate. After 20 h of incubation, the cells in the upper chamber were removed using a cotton stick. Later, the cells that invaded through the Matrigel matrix membrane were stained with 4 μg/ml Calcein AM in Hanks buffered saline at 37°C for 1 h. Each of the experimental conditions was performed in triplicates. Subsequently, fluorescence of the invaded cells was read in an ULTRA Multifunctional Microplate Reader (Tecan) at excitation/emission wavelengths of 530/590 nm. These fluorescently labeled invasive cells were also photographed under a fluorescent microscope.
Protein extraction and western blotting. The PANC-1 and BxPC-3 cell lines were treated with or without OPP for 72 hours to evaluate the effects of treatment on PARP, cleaved caspase-3, cleaved caspase-9, survivin, Bcl-XL and β-actin expressions. Cells were lysed in cold lysis buffer for 30 min on ice. Protein concentrations were determined using the Bradford protein assay kit (Bio-Rad Laboratories, CA, USA). Each sample contained 50μg of total cell lysates. The samples were loaded on 10% SDS-polyacrylamide gels and subjected to electrophoresis. After electrophoresis, the gel was electrophoretically transferred to a nitrocellulose membrane (Whatman,; Clifton, NJ, USA) using transfer buffer (25mM Tris, 190mM glycine, 20% methanol) in a Hoefer TE70XP transfer apparatus (Holliston, MA, USA). The membranes were incubated for 1 hour at room temperature with 5% nonfat dried milk in 1 × TBS buffer containing 0.1% Tween. After that, membranes were incubated overnight at 4°C with primary antibodies (1:1000). The membranes were washed 3 times with TBS-T and subsequently incubated with the secondary antibodies (1:5000) containing 2% BSA for 2 h at room temperature. The signal intensity was then measured by chemiluminescent image with chemiDoc XRS (Bio-Rad Laboratories)
Real-time quantitative PCR for gene expression analysis. Total RNA was isolated using the RNeasy Mini Kit from QIAGEN (Valencia, CA, USA) according to the manufacturer's protocol. Two micrograms of total RNA from each sample were subjected to first strand cDNA synthesis using the TaqMan reverse transcription reagents kit (Applied Biosystems) in a total volume of 20 μl. Reverse transcription reactions were performed at 25°C for 10 min, followed by 48°C for 30 min and 95°C for 5 min. Real-time PCR analysis was performed using the Eppendorf realplex4 system (Hauppauge, NY, USA). The sequences of the primers sets used for this analysis are as follows: MMP-9, forward primer (5’-CGG AGT GAG TTG AAC CAG-3’) and reverse primer (5’-GTC CCA GTG GGG ATT TAC-3’); VEGF, forward primer (5’-GCC TTG CCT TGC TGC TCT AC-3’) and reverse primer (5’-TTC TGC CCTCCT CCT TCT GC-3’); GAPDH, forward primer (5’-CAG TGA GCT TCC CGT TCAG-3’) and reverse primer (5’-ACC CAG AAG ACT GTG GAT GG-3’). All these primers were verified by running them on a virtual PCR and primer concentrations were optimized to avoid primer dimer formation. Real-time PCR amplifications were performed using 2 × SYBR Green PCR Master Mix (Applied Biosystems). Two microliters of RT reaction was used for a total volume of 25-microliter quantitative PCR reactions. The thermal profile for SYBR real-time PCR was 95°C for 10 min followed by 50 cycles of 95°C for 15 s and 60°C for 1 minute. Data were analyzed according to the comparative fold increase or decrease in gene expression determined by quantitation of normalized by GAPDH expression in each sample.
Microwell colorimetric NF-κB assay for measuring NF-κB activity. The TransAM™ Transcription Factor ELISAs kit for P65 (Avtive Motif; Carlsbad, CA, USA) was used to evaluate the binding activity of NF-κB according to the manufacturer's protocol. Briefly, one million of PANC-1 and BxPC-3 cells were seeded in a 100 mm dish. After 24 h incubation, cells were treated with OPP or control for 72 hours. Following that, nuclear protein was extracted from each sample using the nuclear protein extraction kit according to the manufacturer's protocol (Pierce; Rockford, IL, USA). Two micrograms of each sample was incubated in the microplate coated with anti-p65 DNA sequence. In order to detect the p65-DNA binding complex, peroxidase-conjugated anti-DNA antibody was used before color development with ABTS substrate for peroxidase. The chemilumnescence of the samples was determined by using chemiDoc XRS (Bio-Rad Laboratories). The volume of each sample was determined by the Quantity One software (Bio-Rad Laboratories).
Data analysis. Results are expressed as means±SEM and were analyzed using the GraphPad Prism 4.0 (Graph pad Software; La Jolla, CA, USA). Statistical comparisons between groups were done using one-way ANOVA. Values of p<0.05 were considered to be statistically significant and individual p-values are reported in the figures.
Results
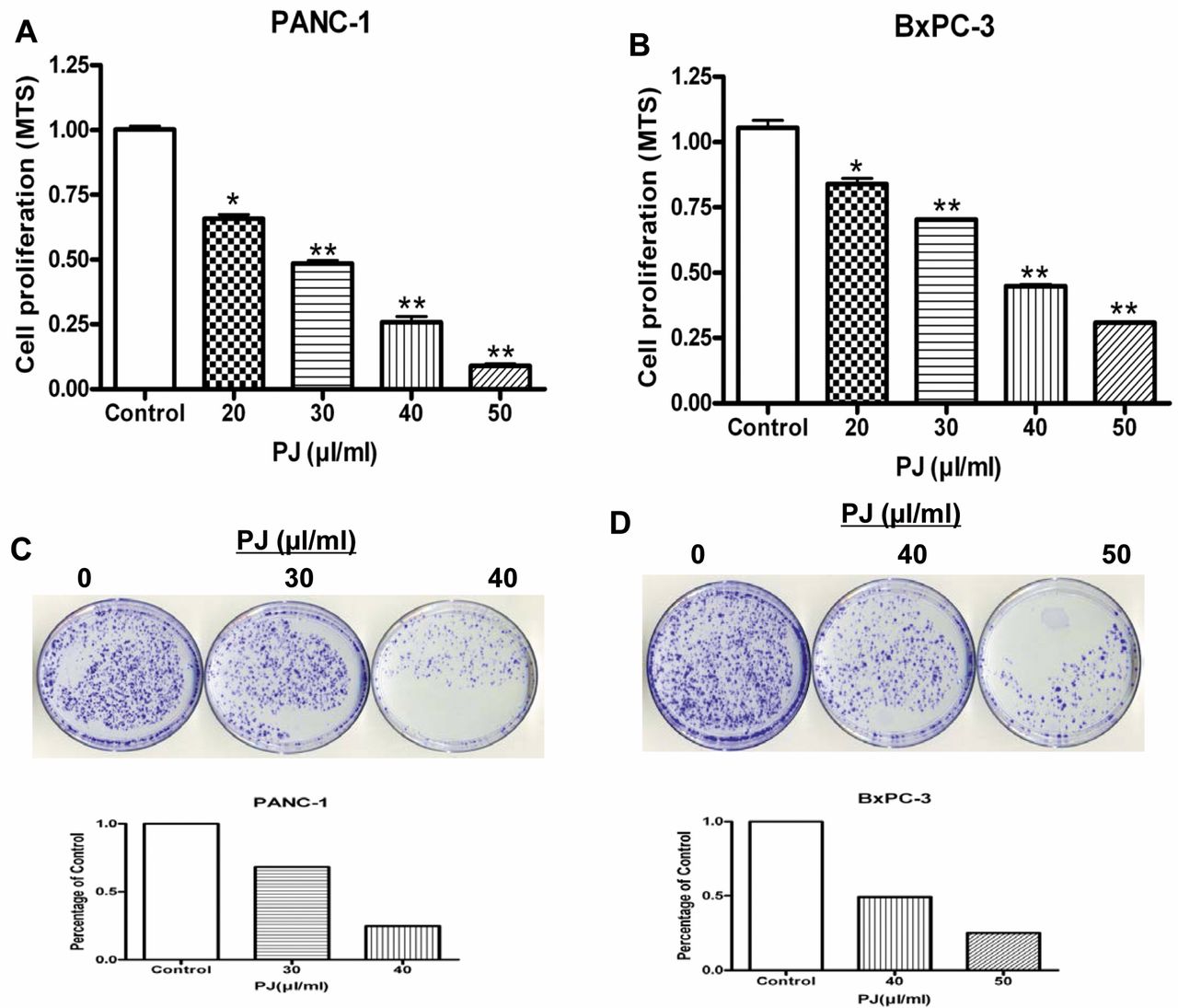
Effects of OPP on cell growth/survival of PaCa cells. To determine the cytotoxic potential of OPP on pancreatic cancer cell lines, PANC-1 and BxPC-3 cells were treated with 20, 30, 40 or 50 μl of OPP (1,500 mg/ml GE) per ml of cell culture medium followed by the MTS and clonogenic assays, as shown in Figure 1A and B. In the PANC-1 cell line, treatment with 20, 30, 40 and 50 μl/ml of OPP for 72 h resulted in 35%, 51%, 75% and 91% inhibition of cell growth relative to control, respectively. Similarly, treatment of the BxPC-3 cell line with 20, 30, 40 and 50 μl/ml of OPP for 72 h resulted in 12%, 30%, 54% and 75% inhibition of cell growth, respectively, relative to control. These results indicate that OPP was an effective inhibitor of pancreatic cancer cell growth as a single-agent. The possibility that the anti-proliferation effect of OPP was due to any alterations in pH was excluded (data not shown).
To confirm the effects of OPP on cell growth, clonogenic assays, as documented in Figure 1C and D, revealed that treatment of PANC-1 and BxPC-3 with increasing concentrations of OPP (30 and 40 μl/ml) resulted in a greater reduction in the number of colonies, as observed by crystal violet staining. Overall, the results from the clonogenic assay were consistent with the MTS data (Figures 1A and 1B), that OPP significantly inhibited pancreatic cancer cell growth.

Effect of OPP (palm juice (PJ)) on pancreatic cancer cell lines PANC-1 and BxPC-3 regarding survival and growth (A and B) and clonogenicity (C and D). Both cancer cell lines were treated with increasing concentrations of OPP. Results are presented as mean±SEM of three assay replicates. *p<0.05 and **p<0.01 versus respective DMSO-treated controls (0 μl OPP). The number of cells counted in the control treatment was considered 100% and the number of cells in OPP-treated cells was calculated relative to this control.
Induction of apoptosis by OPP. The apoptotic ability of OPP was analyzed using the Histone/DNA ELISA method and flow cytometry. As shown in Figure 2A and 2B, OPP induced apoptosis in both cell lines, PANC-1(Figure 2A) and BxPC-3 (Figure 2B) in a dose-dependent manner. Annexin V/PI staining confirmed the apoptosis-inducing effect of OPP in both cell lines tested (Figures 2C and D), respectively. Figures 2C and D represent quantitation of apoptotic cells, as detected by Annexin V staining after treatment with 20 and 30 μl/ml OPP. Our results showed that OPP treatment resulted in a statistically significant (**p<0.01) increase in the percentage of apoptotic cells in both pancreatic cancer cell lines at the IC50 dosage of OPP.
Analysis of cell-cycle distribution after treatment with OPP. To determine possible underlying mechanisms of cell proliferation suppression, cell cycle distribution of PANC-1 (Figure 3A) and BxPC-3 (Figure 3B) cells treated with varying concentrations of OPP were analyzed. After a 72-h incubation period, the distribution of populations in different cell-cycle phases was quantified using flow cytometry. Our results showed that both cell lines were arrested in the S phase by OPP in a dose-depended manner. For PANC-1 cells (Figure 3A), there were about 28% of the cells in S phase in the treatment group (40 μl/ml) compared to 25% in control cells. A similar response was observed in the BxPC-3 cells (Figure 3B) with 52% of cells in S phase in the treatment group (50 μl/ml) compared to 21% in control cells. These findings reveal that OPP induced cell cycle arrest in the S phase for both cancer cell lines.

Apoptosis analysis by the Histone Kit/DNA ELISA method (A and B) and flow cyctometry (C and D). Cells were either vehicle treated (DMSO-control) or treated with increasing concentrations of OPP (palm juice (PJ)) for 72 h and, in addition, were stained with annexin V and propidium iodide for flow cytometry analysis. The flow cytometric cell distribution of cells is illustrated in dot plots (C, D). The values represent±SE of triplicate samples. *p<0.05 and **p<0.01 versus DMSO-treated control groups (0 μl OPP).
OPP up-regulates pro-apoptotic genes and down-regulates anti-apoptotic genes. In order to further understand the molecular mechanisms involved in OPP-induced apoptosis of pancreatic cancer cells, modifications in the cell death pathway were investigated by western blot analysis. Specifically, the expression of proteins involved in the induction of cleaved caspase-dependent apoptosis (caspase-3, caspase-9), anti-apoptotic Bcl-2 family (Bcl-XL) and the anti-apoptotic protein survivin, were analyzed. OPP increased the pro-apoptotic proteins cleaved caspase-3 and - 9 and PARP, in both cancer cell lines. Furthermore, OPP down-regulated the production of the anti-apoptotic proteins such as Bcl-XL and survivin in PANC-1 and BxPC-3 (Figure 4). This analysis demonstrates that OPP induces apoptosis in both cancer cell lines through a dual-mechanistic approach: activation of apoptosis inducing caspases and inhibition of cell survival proteins.
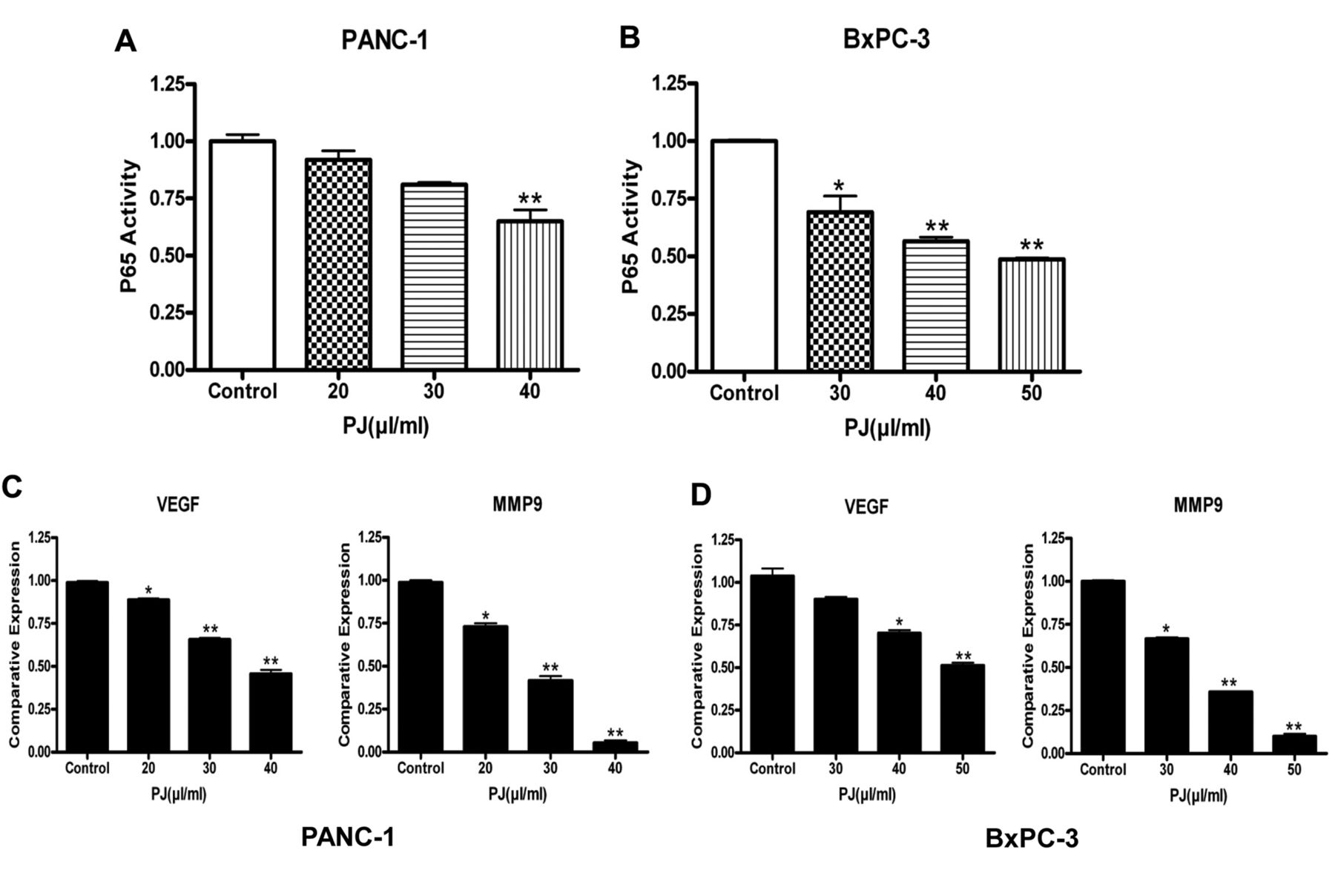
Inhibition of NF-κB activity with OPP. The NF-κB pathway plays important roles in cancer cell transformation, cell invasion and apoptosis. P65 is a subunit of NF-κB that interacts with promoter sequence of target genes that in turn induces the expression of genes involved in inflammation, anti-apoptosis and cell proliferation. To characterize the effect of OPP on NF-κB activity, a microwell colorimetric NF-κB assay was utilized. Our results showed an OPP dose-dependent down-regulatory trend of p65 in both PANC-1 and BxPC-3 cancer cells (Figures 5A and B). Both cell lines (PANC-1 and BxPC-3) exhibited significantly lower NF-κB (p65) activity at higher concentrations of OPP. It is known that vascular endothelial growth factor (VEGF) and matrix metalloproteinase 9 (MMP9) are down-stream genes of NF-κB, responsible for cancer cell invasion and migration expression. The expressions of VEGF and MMP9 were evaluated by real-time PCR after reverse transcriptase PCR. As shown in Figures 5C and D, the expressions of VEGF and MMP9 in both cell lines were significantly inhibited on treatment with OPP. These results strongly suggest that OPP inhibited NF-kB activity and the expression of its target genes.

Response of OPP (palm juice (PJ)) on cell-cycle regulation in PANC-1 (A) and BxPC-3 (B) cancer cell lines. Representative cell-cycle distribution of cancer cells treated with increasing concentrations of OPP (palm juice (PJ)) are shown along with the percentages of the cell-cycle stages, G0-G1, G2-M and S.
Inhibition of cell migration and invasion by OPP. The cell invasion and wound healing assays were utilized to evaluate the effect of OPP on invasion and migration capabilities of the pancreatic cancer cell lines. As depicted in Figure 6A and B, OPP caused a dose-dependent reduction of cell invasion in PANC-1 and BxPC-3. Subsequently, the cells were stained to detect the fluorescence activity of PANC-1 and BxPC-3. The results obtained from this analysis are consistent with the results of the cell invasion assay.
The results obtained from the migration assay are depicted in Figure 6C and D. A dose-dependent inhibition of pancreatic cancer cell migration was observed on analysis of the plates after 20 hours of incubation with various concentrations of OPP. This demonstrates that OPP effectively retards migration and metastasis in PANC-1 and BxPC-3 cells.
Discussion
Pancreatic cancers represent one of the major causes of cancer-related mortality world-wide. The chemotherapeutic agents currently available for pancreatic cancer show highly toxic side-effects and drug resistance, thereby limiting their efficacy and usefulness. Erlotinib, a targeted-drug used as an adjuvant to gemcitabine in pancreatic chemotherapy, works by inhibiting cell growth signaling and has exhibited marginal therapeutic benefits in clinical trials (4). However, side effects of this drug, including heart attack, stroke and even death, have been documented (20). This necessitates the development of new strategies that reduce adverse effects and enhance beneficial output. Thus, nutraceuticals or bioactive components from plants, with their known beneficial pleotropic effects, alone or in combination with drugs, may be an option worth exploring in this endeavor.

Effect of OPP (palm juice (PJ)) on gene expression. Representative western blots showing the effects of increasing palm juice or OPP concentration on the expression of PARP cleavage, caspases, Bcl-xL and survivin in PANC-1 and BxPC-3 cancer cell lines after 72 h. The β-actin protein was utilized as the protein loading control for this experiment.
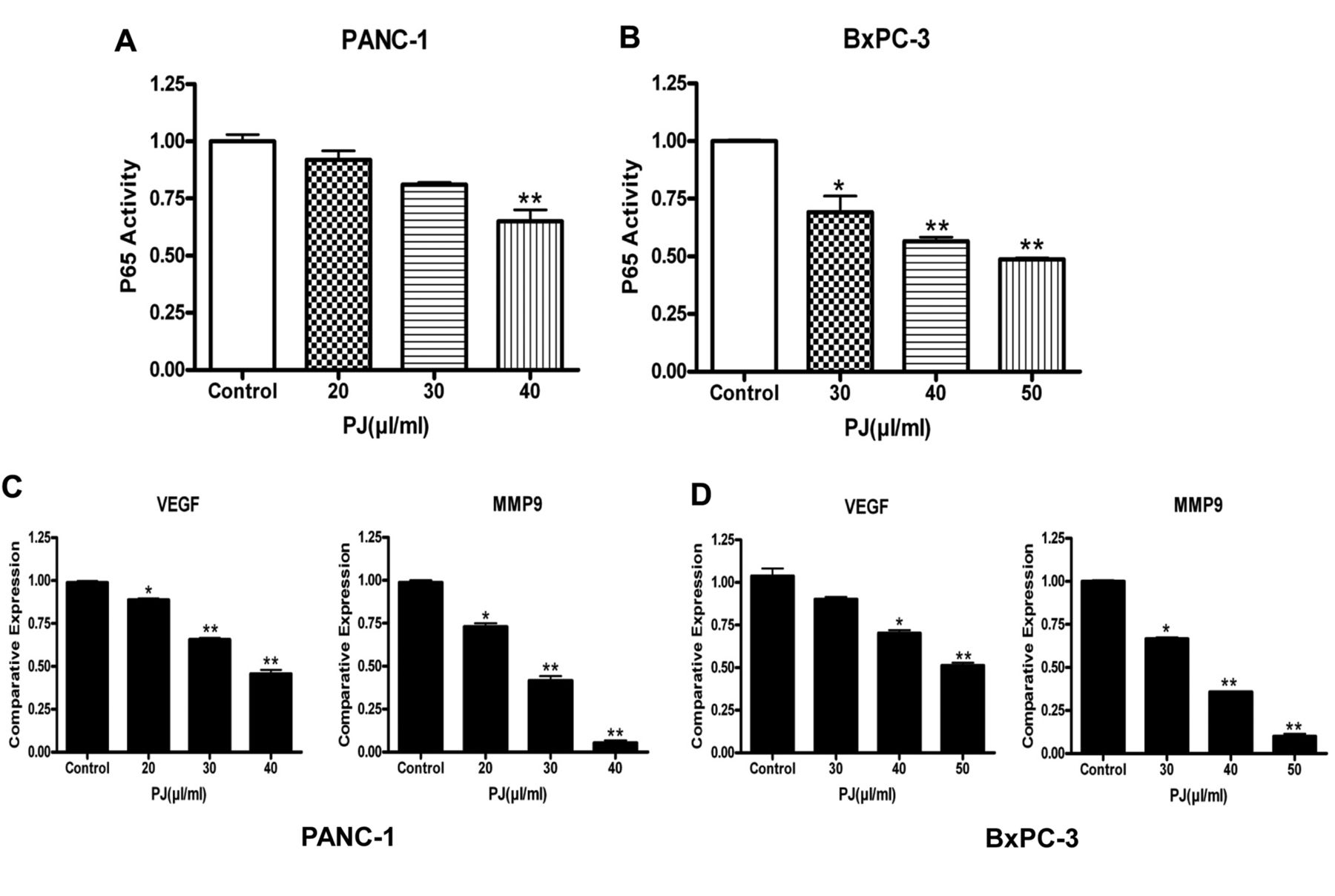
Down-regulation of NFκB activity following OPP (palm juice (PJ)) treatment in PANC-1 and BxPC-3 cells. A and B: Analysis of p65 activity in PANC-1 and BxPC-3. P65 activation decreased in a dose-dependent manner following OPP (palm juice (PJ)) treatment in both cell lines. C, D: Effect of OPP on VEGF and MMP-9 expression relative to the DMSO treated control. OPP decreased VEGF and MMP-9 expression in a dose-dependent manner. The effect on MMP-9 was more pronounced in both cancer cell lines. Results are presented as mean±SEM of three assay replicates. *p<0.05 and **p<0.01 versus respective DMSO treated controls.

OPP (palm juice (PJ)) inhibits cell invasion and migration in PANC-1 and BxPC-3 cell lines. The migration of PANC-1 (A) and BxPC-3 (B) cells was evaluated using a cell invasion assay. Cells were cultured in the absence of, or in varying concentrations of, OPP for 20 h and were photographed after staining with Calcein AM. Then, fluorescence of the invaded cells was determined (A, B). OPP decreased cell invasion in a dose-dependent manner, as semi-quantified by the fluorescence index reading. Results are presented as mean±SEM of three assay replicates. *p<0.05 and **p<0.01 versus respective DMSO treated controls (0 μl OPP). Further evaluation of cell migration was done using the wound healing assay (C, D). When the cells reached confluence, a single wound was scratched across each well. After a-20h incubation period, wound closure areas were visualized using a microscope. These results also support the cell invasion findings that increasing OPP concentrations prevent cell migration in an increased manner.
Palm Juice (PJ) or OPP, a water soluble extract obtained from the oil palm fruit, rich in phenolics, has been shown to have potential chemotherapeutic activity (anti-inflammatory, inhibition of cell growth and proliferation, pro-apoptotic and anti-oxidative) in vitro (17). We investigated the effects of OPP in the pancreatic cancer cell lines PANC-1 and BxPC-3. Our results show that OPP inhibits proliferation, growth, migration and invasion of both cell lines by inducing apoptosis, cell cycle arrest and down-regulating NF-κB activity.
To determine the effectiveness of OPP against cancer cell growth and survival, we performed the MTS and colonogenic assays. We found that, while OPP inhibited cell proliferation and clonogenicity in both cell lines, PANC-1 cells were more sensitive to the effects of OPP than BxPC-3 cells for all concentrations tested (Figure 1). This may be attributed to the genetic differences in the two cell lines including the K-RAS mutation being present in the PANC-1 cells, as opposed to the BxPC3 cell line, which has a wild type K-RAS gene. This is of specific interest since most pancreatic cancer patients carrying a mutated K-RAS have a less favorable prognosis on current therapeutics (21). In order to analyze whether these results were due to the induction of apoptosis, the histone/DNA ELISA assay and flow cytometry to test for apoptosis induced by OPP were performed. OPP was effective in inducing apoptosis in a dose-dependent manner for PANC-1 and BxPC-3 (Figure 2). Furthermore, the flow cytometry analysis on cell-cycle distribution revealed that OPP induces cell-cycle arrest in the S phase for both PANC-1 and BxPC-3 cells (Figure 3). Taken together, these data suggest that OPP inhibits cell growth and induces apoptosis with the ability to induce cell-cycle arrest in these cancer cell lines and, hence, could potentially prove to be an effective antitumor agent.
Since the activation of caspases and PARP protein are important factors involved in various apoptotic pathways, we also evaluated the expressions of these proteins by western blot analysis. Additionally, we investigated whether OPP also simultaneously caused a decrease in the expressions of the anti-apoptotic proteins such as Bcl-xL and survivin in both PANC-1 and BxPC-3 cell lines. Our data demonstrated that OPP can promote the expressions of pro-apoptotic proteins, such as cleaved caspase-3, cleaved caspase-9 and cleaved PARP. Additionally, OPP can reduce the anti-apoptosis protein expression including Bcl-XL and survivin in both cancer cell lines and this beneficial effect was observed to be dose-dependent (Figure 4). Hence, our current study revealed that the apoptotic capacity of OPP is induced through a dual mechanism, demonstrating that OPP can indeed be a powerful nutraceutical against cancer.
NF-κB plays important roles in many cellular processes including cell proliferation, anti-apoptosis invasion and angiogenesis, all of which are crucial for cancer development and progression (6). Our data demonstrated that P65 (a subunit of NF-κB) activity significantly decreased as OPP concentration increased in both cell lines (Figure 5A and B). Since VEGF and MMP9 are known to be involved in downstream signaling of NF-κB responsible for cell migration and invasion, we evaluated VEGF and MMP9 relative expressions for both cell lines. Our results confirm that expression of both VEGF and MMP9 was significantly decreased in response to increased OPP concentrations in both PANC-1 and BxPC-3 cell lines (Figure 5C and D).
To verify OPP exhibited properties to inhibit cell invasion and migration, we carried-out cell invasion and wound healing assays. Our results show that OPP caused a dose-dependent reduction in cell invasion and migration (via the wound healing assay) in both PANC-1 and BxPC-3 cells (Figure 6). This further supports OPP's candidacy in treating cancer.
In summary, our data demonstrate the potential for palm juice (PJ) or OPP as a multi-action chemotherapeutic agent against pancreatic cancer. Benefits of this nutraceutical involve inhibition of cell proliferation and growth and induction of cellcycle arrest and apoptosis. The mechanisms through which these actions occur have also been partially elucidated in this study, the most important of which is the evident down-regulation of the NF-κB pathway. However, further investigation in animal models should be conducted in order to fully gauge its potential in the prevention and treatment of pancreatic cancer.
Acknowledgements
This work was funded in part by the Malaysian Palm Oil Board (MPOB), Kuala Lumpur, Malaysia.
Footnotes
-
This article is freely accessible online.
- Received August 13, 2014.
- Revision received September 24, 2014.
- Accepted September 29, 2014.
- Copyright© 2015 International Institute of Anticancer Research (Dr. John G. Delinassios), All rights reserved
References
In this issue





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Jump to section
Related Articles
Cited By...
- No citing articles found.