


Abstract
Aim: Tumor angiogenesis is a focus of molecularly-targeted therapies. This study investigated the effect of sodium butyrate (SB), a histone deacetylase inhibitor, on the synthesis of antiangiogenic and lymphangiogenic factors in oral squamous cell carcinoma. Design: Gene alterations in HSC-3 cells were assessed using cDNA microarrays before and after treatment with SB. The mRNA and protein expression of lymphangiogenic factors were also assessed by quantitative PCR, western blotting and immunocytochemistry. Results: Microarray analysis revealed that treatment with SB led to altered expression of angiogenesis-related gene expression. The quantitative polymerase chain reaction showed that platelet-derived growth factor-B, angiopoietin-2, vascular endothelial growth factor (VEGF)-C, and VEGFD were down-regulated. Western blotting and immunocytochemistry confirmed reduced protein synthesis of VEGFC. Conclusion: SB inhibits expression of lymphangiogenic factors in HSC-3 cells. Within the limitations of the present study, SB may have potential as an anti-metastatic pro-drug for oral cancer.
The metastasis of tumors to regional lymph nodes and distant organs is a major cause of cancer-related deaths. During metastasis, tumor cells invade into microvessels close to the tumor nests, and the density of lymphatic vessels around the nests is therefore an important factor. Recent studies suggest that lymphangiogenic factors are secreted directly from tumor cells to induce lymph vessels to facilitate sentinel lymph node metastasis (1). Families of growth factors such as vascular endothelial growth factor (VEGFA, -B, -C and -D), angiopoietin-2 (ANGPT2) and platelet-derived endothelial growth factor (PDGF) are secreted from tumor cells and play roles in both angiogenesis and lymphangiogenesis (2). VEGFC was first-identified as a lymphangiogenic factor, and its expression increases in primary tumors of various human carcinomas. The expression of VEGFC in tumor cells was also correlated with the frequency of tumor metastasis to regional lymph nodes (1, 3). VEGFC binds to VEGF tyrosine kinase receptor-2 and -3 (VEGFR2 and -3) in endothelial cells to induce proliferation and migration as well as an increase in vascular permeability. Although VEGFD has a cysteine-rich-COOH terminal extension, its structure is similar to that of VEGFC. VEGFD also binds to the VEGFR2 and -3, and may also play a role in lymphangiogenesis. These data suggest that VEGFC and -D are potential molecular targets for the treatment of malignant tumors (4-7).
Gene transcription is strictly controlled by a variety of mechanisms. Recently, novel cancer therapies have focused on the epigenetic control of gene transcription. The acetylation and a de-acetylation of histones regulate gene expression by modulating the conformation of chromatin. Therefore, histone deacetylase (HDAC) inhibitors have emerged as potential anticancer agents (8-10). Sodium butyrate (SB), a four-carbon fatty acid found abundantly in milk fat, is also produced by the microbial fermentation of dietary fibre in the colonic lumen of mammals. SB is a HDAC inhibitor that induces cell-cycle arrest, differentiation and apoptosis of oral cancer cells (11-14). In a previous study, we found that SB inhibited the growth of oral cancer cells in a dose- and time-dependent manner. Treating the human oral squamous cell carcinoma (OSCC) cell line HSC-3 with SB induces cell-cycle arrest in the G1 phase by increasing the transcription of cyclin-dependent kinase inhibitor p21WAF1 and phospholipase A2 (13). SB also inhibits proliferation and induces the differentiation and apoptosis of a variety of tumor cell types, including erythroleukaemia, embryonal carcinoma, colonic carcinoma, pancreatic carcinoma, neuroblastoma, glioblastoma and haematopoietic malignancies (15).
To assess effects of SB on lymphangiogenesis, we examined the alterations in the transcription of VEGFA, VEGFC, VEGFD and ANGPT2 in the OSCC cell line HSC-3 using cDNA microarrays, quantitative polymerase chain reaction (PCR), immunocytochemistry and western blotting.
Materials and Methods
Cell culture and incubation with SB. The human OSCC cell line HSC-3 was obtained from the Japanese Cancer Research Resources Bank (Tsukuba, Japan) and maintained in RPMI-1640 medium supplemented with 10% foetal bovine serum and 1% penicillin/streptomycin (Gibco BRL, Grand Island, NY, USA) at 37°C with 5% CO2 until they reached 75-90% confluence. Cells were then washed twice with phosphate-buffered saline (PBS), incubated in culture medium with or without SB (0, 1, 3 mM; Wako Pure Chemical Industries, Ltd., Osaka, Japan) and analysed.
Total RNA extraction. Total RNA was isolated from HSC-3 cells using an RNeasy Mini Kit (Qiagen, Inc., Valencia, CA, USA) following the manufacturer's instructions, and the quantity and purity were assessed using standard spectrophotometric methods.
cDNA microarray analyses. For gene expression profiling, we used an Affymetrix GeneChip® Human Genome Focus Array (Affymetrix, Santa Clara, CA, USA) following the manufacturer's instructions. Briefly, 2×106 cells were seeded in a 25-cm2 flask. After 48 h, cells were incubated with SB (0, 1, 3 mM) for 6 or 12 h, as indicated. Total RNA was extracted and mixed with T7-oligo (dT) promoter primer, reverse-transcribed into cDNA and then into double-stranded cDNA using a GeneChip® Expression 3’-Amplification Reagents Two-cycle cDNA Synthesis Kit (Affymetrix). The double-stranded cDNA was purified and used as a template for in vitro transcription (IVT) using a GeneChip® IVT Labelling Kit (Affymetrix). The biotinylated cRNA target was then fragmented and hybridised to a GeneChip® Human Focus Array (Affymetrix), which contained a broad range of genes derived from publicly available well-annotated mRNA sequences. Immediately after hybridisation, the array was washed and stained with streptavidin–phycoerythrin (Molecular Probes, Eugene, OR, USA). Each probe array was scanned twice; the average of the two images was calculated and the intensity under each condition was compared. Differentially expressed genes were defined as having a greater than twofold variation in gene expression. Gene ontology was classified according to the Gene Ontology Consortium criteria using GeneSpring software (Silicon Genetics, Redwood City, CA, USA).
Reverse transcription and quantitative PCR. cDNAs were synthesised using a PrimeScript RT Reagent Kit (Takara Bio Inc., Shiga, Japan) according to the manufacturer's instructions. Quantitative PCR was performed using 2 μl template cDNA and 23 μl PCR mixture (SYBR Premix Ex Taq; Takara Bio Inc.) including 10 pmol each of forward and reverse primers. The primers were selected from the highly specific sequence of each gene (Table I). Amplification was performed using a Smart Cycler II system (Cepheid, Sunnyvale, CA, USA), with preheating at 95°C for 1 min, followed by 40 cycles at 94°C for 30 s and 60°C for 30 s. Gene transcription levels were normalised to these of glyceraldehyde 3-phosphate dehydrogenase (GAPDH), and data from SB-treated cells were expressed as fold induction compared with untreated control cells. The levels of GAPDH transcription under each condition were confirmed to be unchanged. All samples were analysed in triplicate.
Immunocytochemistry. HSC-3 cells were seeded on 22×24 mm cover glasses (C022241; Matsunami Glass Ind., Ltd., Osaka, Japan), in ϕ35-mm culture dishes and incubated in media in the presence or absence of 3 mM SB for 24 h. After incubation, cells were washed three times with Tris-buffered saline (TBS), fixed in ice-cold 10% buffered formalin for 10 min and then washed again with TBS. Haematoxylin and eosin (H/E) staining was then performed using conventional methods. Immunocytochemistry for VEGFA and VEGFC was then carried out. Antigen retrieval was performed by incubating the samples in 10 mM citrate buffer (pH 6.0) at 98°C for 20 min. Samples were incubated for 5 min at room temperature with 1% bovine serum albumin (BSA) containing 0.1% Tween 20 (TBS-T). They were then incubated with primary antibodies for 60 min at room temperature as follows: VEGFA (sc-152, 1:100; Santa Cruz Biotechnology, Inc., Santa Cruz, CA, USA), VEGFC (18-2255, 1:100; Zymed Laboratories Inc., South San Francisco, CA, USA). For the negative control, primary antibodies were replaced with 1% BSA/TBS-T. Endogenous peroxidase activity was quenched using 3% hydrogen peroxide. Cells were then incubated with an Envision/horseradish peroxidase (HRP) universal kit (K1390; Dako, Glostrup, Denmark) for 60 min at room temperature, and the immune complexes were visualised using 3,3’-diaminobenzidine tetrahydrochloride as the chromogenic substrate. Finally, the specimens were lightly counterstained with Mayer's haematoxylin, dehydrated in a graded ethanol series and mounted with malinol (Wako Pure Chemical Industries, Ltd.).
Immunostaining was considered to be positive when at least moderate staining was detected in the cytoplasm. Immunoreactivity was evaluated using a semiquantitative scoring system: (0, no staining; 1+, <50% positive staining; 2+, 50-75% positive staining; 3+, >75% positive staining).
Western blotting. HSC-3 cells were treated with SB (3 mM) for 24 h. After incubation, cells were washed three times with PBS and lysed in buffer containing 50 mM Tris–HCl (pH 7.5), 150 mM NaCl and 1% Triton X-100 supplemented with a 1/100 (vol/vol) protease inhibitor cocktail (P8340; Sigma-Aldrich Co., St. Louis, MO, USA). Cell lysate was then centrifuged at 21,000× g for 5 min at 4°C to remove cellular debris. The protein concentrations in the cell lysates were then determined using a Bio-Rad Protein Assay (Bio-Rad Laboratories Japan, Tokyo, Japan). Eighty micrograms of each sample was then separated by sodium dodecyl sulphate-polyacrylamide gel electrophoresis (SDS-PAGE) on 10% gels and transferred to polyvinylidene difluoride (PVDF) membranes (Immobilon-P; Millipore Corporate, Billerica, MA, USA). Membranes were then blocked using blocking reagent (NYPBR01; Toyobo, Osaka, Japan) and incubated overnight with antibodies against VEGFA (1:1000; Santa Cruz Biotechnology Inc.) or VEGFC (1:1000; Cell Signaling Technology, Inc., Danvers, MA, USA). After washing with TBS-T, the membranes were incubated with horseradish peroxidase-conjugated goat anti-rabbit IgG secondary antibodies (1:20,000; Santa Cruz Biotechnology Inc.) for 1 h at room temperature. The reaction products were then visualised using the ECL plus detection system (GE Healthcare Life Sciences, Little Chalfont, Buckinghamshire, UK). Membranes were exposed to Kodak Biomax film (GE Healthcare Biosciences) and then re-blotted with an antibody against β-actin (ab-8226, 1:2,000; Abcam, Cambridge, UK). The membranes were washed three times for 15 min with TBS-T between each step.
Primers used in quantitative polymerase chain reaction.
Statistical analysis. Statistical significance of differences was determined using a Kruskal–Wallis H-test. Values of p<0.05 were considered to be statistically significant.
Results
Transcription of angiogenic- and lymphangiogenic-related genes. To investigate the effects of SB treatment on gene expression, HSC-3 cells were incubated for 6 or 12 h in the presence or absence of SB (1 and 3 mM). cDNA microarray analysis identified 1,925 down-regulated and 4,381 up-regulated genes in SB-treated HSC-3 cells compared with the control. Down-regulated genes included those encoding proteins related to angiogenesis and lymphangiogenesis such as platelet-derived growth factor B (PDGFB) VEGFA, VEGFB and VEGFC, as shown in Table II. These data were confirmed by quantitative PCR. The transcription of angiogenic factors such as ANGPT2 was suppressed in a dose-dependent manner at 3 h after incubation with SB (Figure 1). Furthermore, the transcription of PDGFB was down-regulated as early as 30 min and remained low, but not statistically significant. The expression levels of VEGFA and VEGFC were not altered until 1 h. The expression of VEGFA increased at 3 h, but not significantly, in contrast to VEGFC, which decreased significantly at 3 h after incubation of cells with SB. The expression of VEGFD decreased significantly at 30 min and remained low. Finally, the expression of cyclooxygenase 2 (COX2) increased after 30 min and continued to rise at later time points.
Alterations in the translation levels of angiogenic and lymphangiogenic genes related to HSC-3 cells following treatment with sodium butyrate (SB). Transcription levels are expressed as the ratio of SB-treated groups compared with the untreated control. Data represented as having a greater than twofold variation from the control.
Effects of sodium butyrate (SB) on expression of vascular endothelial growth factor (VEGFA, -C) in HSC-3 cells.
Western blotting was used to confirm the changes in expression of VEGFA and VEGFC at the protein level in SB-treated HSC-3 cells. VEGFA was not detected in untreated cells, but low levels were detected after 3 h of treatment with SB. In contrast, VEGFC was clearly detected in untreated cells, expression of which decreased following treatment with SB (Figure 2). These findings were consistent with our quantitative PCR data. The cytoplasmic expression of VEGFA and VEGFB remained unchanged after treatment with SB (data not shown). Furthermore, these cells had no evidence of apoptotic bodies after SB treatment.
Immunostaining was used to investigate the expression of VEGFA and VEGFC in HSC-3 cells (Figure 3). VEGFA-positive staining was apparent in a fine granular form around the nucleus, and the staining intensity increased slightly on treatment with 3 mM of SB. In contrast, high expression of VEGFC was detected in untreated cells, this clearly decreased after treatment with 3 mM of SB (Figure 3, Table III). These immunostaining results are consistent with quantitative PCR and western blotting data.
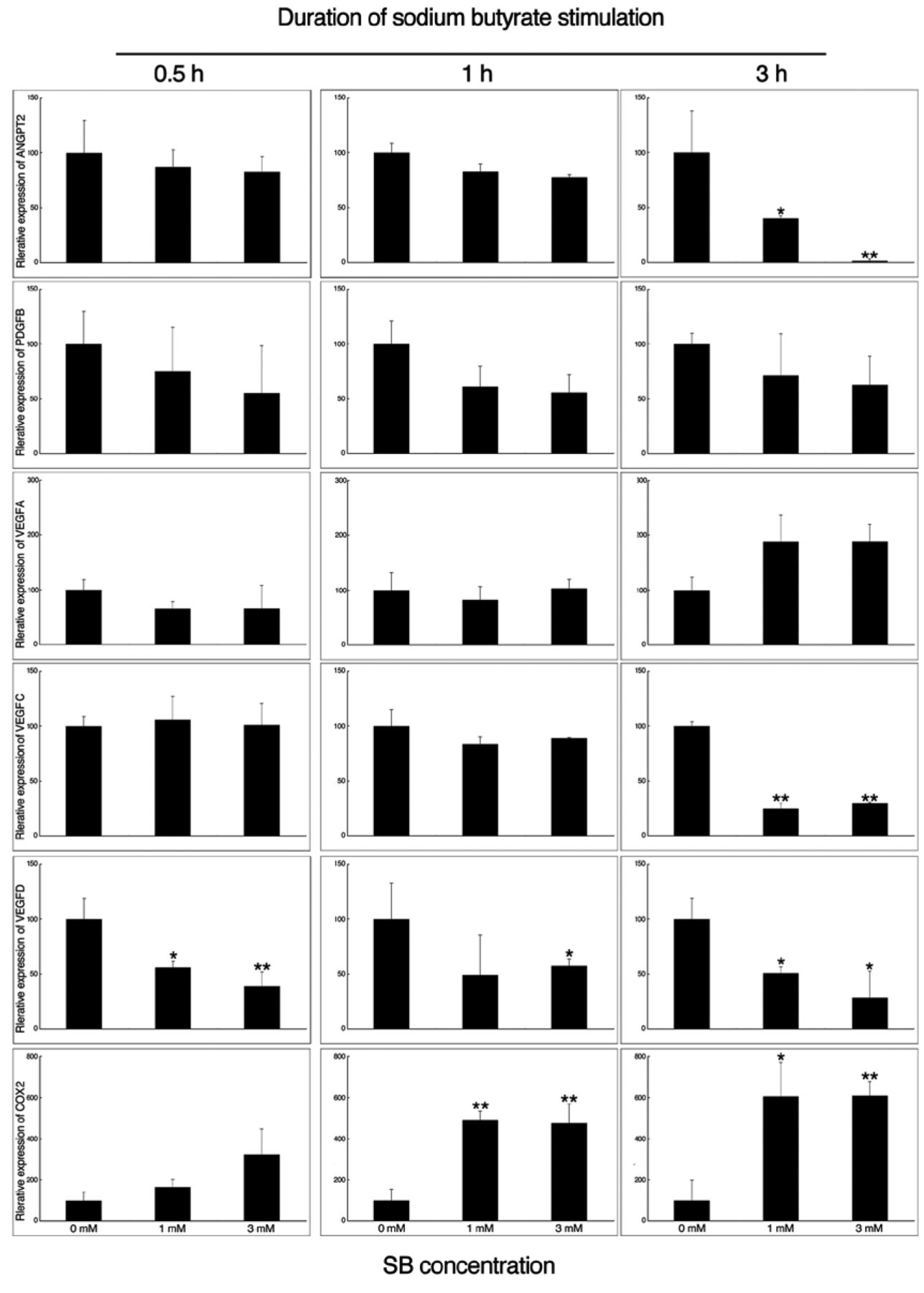
Alterations in the transcription of angiogenic- and lymphangiogenic-related genes. HSC-3 cells were incubated with 0, 1 or 3 mM of butyrate (SB) and analyzed by PCR. The mRNA expression of ANGP2, PDGFB, VEGFD and VEGFD were down-regulated by SB. In contrast, the expression of VEGFA and cyclo-oxygenase COX2 increased. GAPDH was used as internal control. *p<0.05, **p<0.01 compared to the untreated control.

Western blotting of vascular endothelial cell growth factor (VEGF)-A, VEGFC and β-actin in HSC-3 cells. HSC-3 cells were incubated with or without sodium butyrate (SB) for 24 h and analyzed by western blotting. The expression of VEGFA increased following SB incubation, whereas the expression of VEGFC decreased.
Discussion
The present study demonstrated that treatment with SB modulated transcription in the human OSCC cell line HSC-3. Our cDNA microarray analysis demonstrated that treatment with SB resulted in down-regulation of genes related to angiogenesis and lymphangiogenesis namely ANGPT2, PDGFB, VEGFC, and VEGFD. Out of these, VEGFC is of particular interest because its expression was correlated with lymphatic vessel density and lymph node metastasis in OSCC (16). Consistent with this finding, VEGFC expression was detected in oral carcinoma of the tongue using immunohistochemistry (1).
VEGFC plays pivotal roles in lymphangiogenesis. Overexpressing VEGFC in the non-VEGFC-expressing and non-metastatic human breast cancer cell line MCF-7 resulted in enhanced tumor growth, and lymphangiogenesis and lymphatic metastasis in immunodeficient mice (17). The increased expression of VEGFC in tumor tissues results in a poor prognosis and is positively correlated with lymph node metastasis in many types of cancers including those of breast (18), uterine cervix (19), colon and rectum (20), oesophagus (21), stomach (22), head and neck (23), and gallbladder (24). In addition, serum VEGFC was elevated in patients with non-small cell lung cancer (25), and colorectal cancer (26). In non-small cell lung cancer, this was also correlated with lymph node metastasis. VEGFC promotes lymphangiogenesis by binding to vascular endothelial cell growth factor receptor 3 (VEGFR3) on lymphatic endothelial cells. VEGFD is another member of the VEGF family that can bind to VEGFR3 and exert effects on lymphangiogenesis (27). In vitro data revealed that VEGFD is the most potent angiogenic and lymphangiogenic effector in the VEGF family (7). Our data suggest that SB also down-regulates expression of VEGFD. This might contribute to inhibition of lymphangiogenesis in cooperation with VEGFC.

Altered cytoplasmic expression of vascular endothelial cell growth factor (VEGF)-A and VEGFC in HSC-3 cells. HSC-3 cells were incubated with or without sodium butyrate (SB) on cover slip for 24 h. Treatment with SB induced cytoplasmic VEGFA expression, whereas VEGFC expression was markedly down-regulated. Scale bar=10 mm.
Solid tumor growth cannot exceed 1-2 mm3 without an adequate blood supply, and so angiogenesis is a crucial step in tumor growth, invasion and metastasis (28). Moreover, tumor angiogenesis is associated with tumor progression and aggressiveness in various malignancies (29, 30), including head and neck carcinomas (31). EGF expression was detected in OSCC (32), and so the EGF family may play a central role in tumor growth and metastasis. However, the expression of VEGFs in OSCC has not been fully elucidated.
VEGFA may also promote lymphatic vessel formation. Injection of an adenoviral vector expressing VEGFA into the ears of mice induced formation dilated lymphatic vessels (34). Supplementation with VEGFA promoted the sprouting of lymphatic vessels from pre-existing blood vessels in embryoid bodies (34). Lymphangiogenesis was also observed in mice with skin SCCs and SCC metastasis-positive lymph nodes (35). In contrast, VEGFA-containing corneal implants or the injection of a VEGFA-expressing recombinant adenoviral vector in skeletal muscle did not induce lymphangiogenesis (7, 36, 37). Therefore, the role of VEGFA in lymphangiogenesis is controversial, and further studies are required to clarify its effects.
Butyrate inhibits the growth and proliferation of a variety of tumor cells including erythroleukaemia, embryonal, colon carcinoma, pancreatic carcinoma, neuroblastoma, glioblastoma and haematopoietic malignancies (15). We previously showed that SB inhibited the proliferation of HSC-3 cells and increased the expression of phospholipase-A2-X and COX2 (13). The effects of SB on transcriptional regulation and post-transcriptional modifications have been reported. Specifically, SB was proposed to non-competitively inhibit HDACs, which results in the modulation of gene transcription. SB-induced cell growth inhibition may be due to the increased transcription of cyclin-dependent kinase inhibitor p21KIP/WAF, which leads to cell cycle arrest in the G2 phase (13). Recently, HDAC inhibitors including SB, trichostatin A, LAQ824, FK228 and sulphoraphane, were reported to down-regulate hypoxia inducible factor (HIF)-1α in cancer cell lines (38-42). The expression of GATA binding protein 2 and VEGF family members are also strongly induced under hypoxic conditions via HIF regulated elements (43, 44). The constitutive degradation of HIF1α is blocked during hypoxia due to the oxygen requirements of HIF prolyl-hydroxylases. Heterodimers of HIF1α and HIF-1β are termed the aryl hydrocarbon nuclear translocator. These complexes bind to hypoxia-responsive elements in the promoters of hypoxia-inducible genes and initiate the transcription of more than 100 genes including COX2, metalloproteinase 2 (MMP2), VEGF and VEGFR1 (36, 43, 45). The overexpression of COX2 induces the synthesis of prostaglandin E2 and activates the prostaglandin receptor EP1/EP4. Activation of this receptor induces the tyrosine phosphorylation of human epidermal growth factor receptor HER-2/neu. Signals from HER-2/neu are mediated through mitogen-activated protein kinase p38 and nuclear factor-κB to increase the transcription of VEGF family members (46, 47). However, COX2 and its derived prostaglandins were reported to regulate angiogenesis and lymphangiogenesis via distinct mechanisms, since the functions of COX2 in angiogenesis are mediated predominantly through the prostaglandin receptor EP2 (48, 49). The detailed mechanism by which the transcription of VEGF family members is activated by COX2 remains unclear. Because SB regulates various genes, it may activate an inhibitory cascade involving the expression of ANGPT2, VEGFC and VEGFD.
Inhibition of ANGPT2, VEGFC and VEGFD production by SB may suppress angiogenesis or lymphangiogenesis, suggesting that SB may have therapeutic potential in cancer.
Acknowledgements
This work was supported by Nihon University Sato Grant for 2013 (to Dr Komiyama) and a grant from Dental Research Center, Nihon University School of Dentistry. The works was also supported by Grant-in-Aid for Scientific Research (B) #2379215 and (C) #24593066.
Footnotes
-
This article is freely accessible online.
-
Conflicts of Interest
None.
- Received December 7, 2013.
- Revision received December 25, 2013.
- Accepted December 31, 2013.
- Copyright© 2014 International Institute of Anticancer Research (Dr. John G. Delinassios), All rights reserved
References
In this issue





{kind=link}
{kind=link}
{kind=link}
Jump to section
Related Articles
Cited By...
- No citing articles found.