


Abstract
Background: TNF-related apoptosis-inducing ligand (TRAIL) is considered as a tumor cell-specific cytotoxic agent. Through the aid of mesenchymal stem cells (MSCs), TRAIL is capable of inducing apoptosis of tumor cells in tumor sites. The present study was performed to investigate the cytotoxic effects of TRAIL-engineered MSCs on glioblastoma cells (C6) in vitro. Materials and Methods: An expression vector of secreting form of TRAIL was used to engineer MSCs. The cytotoxic effects of TRAIL-transfected MSCs on C6 cells were invstigated using the MTT method and Hochest33258 staining after co-culture of the two cell types. Results: TRAIL and control plasmid transfection of MSCs showed no significant effect on MSC's viability (p>0.05). A significant inhibition of C6 cells was observed when they were co-cultured with TRAIL-engineered MSCs (63.7%±0.12, p<0.05). Conclusion: Mesenchymal stem cells were very well tolerant to the transfection of TRAIL-bearing vectors. The cytotoxic effects of TRAIL-engineered MSCs on C6 cells indicates the therapeutic potential of this strategy for treatment of glioblastoma patients.
Gliomas are the most common type of primary brain tumors and are often fast-growing with poor prognosis for the patient. The incidence of primary brain tumors worldwide is approximately 7 per 100,000 individuals per year, accounting for 2% of all primary tumors (1). Their complex cellular composition, diffuse invasiveness and capacity to escape conventional therapies have challenged researchers and hampered progress towards an effective treatment over the past three decades (2, 3). To a significant extent, current conventional cancer therapies are symptomatic and passive in nature. The major obstacle to the development of effective cancer therapy is believed to be the absence of sufficient tumor specificity. Since the discovery of tumor-oriented homing capacity of mesenchymal stem cells (MSCs), the application of specific anticancer gene-engineered MSCs has held great potential for the treatment of malignant gliomas (4, 5). MSC-mediated anticancer therapy relies on tumor-specific selectivity provided by MSCs and MSC-carried anticancer agents (6). Homed directly at the tumor microenvironment, engineered MSCs are able to express and/or release anticancer agents to constantly act on the adjacent tumor cells.
TRAIL is a member of the TNF super-family. It was originally identified and cloned on the basis of its sequence homology to the extracellular domain of CD95 ligand (CD95L) and TNF (7, 8). TRAIL is one of few anticancer proteins which selectively causes apoptosis of tumor cells through activation of death receptors, with no effects on healthy cells (9). The ability of TRAIL to selectively induce apoptosis in cancer cells while sparing normal cells makes it an attractive target for the development of new cancer therapies. So far, in this research realm two major areas have been extensively investigated: direct application of recombinant TRAIL or agonistic anti-human TRAILR receptor mABs (monoclonal antibodies), and MSC-based TRAIL engineering. Although pre-clinical studies using recombinant TRAIL (rhTRAIL) have provided evidence for exogenous TRAIL efficacy in suppressing tumor growth, both in vitro and in vivo (10-12), this kind of application is unable to focus solely on targeted cells, thereby systemic side-effects occur, especially in the liver and kidney. However, TRAIL-engineered MSCs are able to dual-target tumor cells and exert anticancer activity efficiently and constantly in tumor microenvironments. As a component part of developing a dual-targeted therapeutic strategy to treat glioma, this study was performed to assess the effects of TRAIL transfection on MSCs and the effects of TRAIL-engineered MSCs on glioma cells.
Primer sequences and PCR product size.
Materials and Methods
Cells and culture conditions. Bone marrow-derived and GFP-transgenic rat MSCs were purchased from Cyagen Biosciences (Guangzhou, China). The GFP-MSCs were cultured in DMEM with 10% FCS, 2 mM L-glutamine and 1% penicillin-streptomycin solution (all from Invitrogen, Shanghai, China) and incubated at 37°C in a humidified atmosphere with 5% CO2. The cells used in this study were limited to passages 9 to 12. The rat C6 glioma cell line (CCL-107) was purchased from the American Type Culture Collection (ATCC, Manassas, VA, USA). C6 cells were maintained as suggested by ATCC and their culture condition was kept consistent with the MSCs. The cells used in the present study were limited to 3 passages.
TRAIL-bearing plasmid transfection of MSCs. A secreting form of TRAIL plasmid (EF030546, gift from the Institute of Life Science, Taihe Hospital, Shiyan, China) was transfected into MSCs with Lipofectamine 2000 (Invitrogen, Shanghai, China), as suggested by the manufacturer. Briefly, the MSCs were plated at 6×105 per well in six-well plates in 3 ml of DMEM medium and dated as day 0. On day 1, the cells were transfected with 2.5 μg of TRAIL plasmid DNA (each well). The control cells were transfected with the same vector without TRAIL insertion.
Reverse transcriptase polymerase chain reaction (RT-PCR). RT-PCR was used to detect the expression of TRAIL on transfected MSCs. The related primers for TRAIL and β–actin are listed in Table I. Total RNA was isolated from MSCs with TRIzol (Invitrogen) 24 h post-transfection, and cDNA was synthesized in the presence of reverse transcriptase at 42°C for 40 min. In the presence of Taq enzyme, PCR was performed 30 circles and each circle was composed of 94°C for 30 sec, 55°C for 30 sec and 72°C for 1 min.
Immunofluorescent detection of TRAIL expression in MSCs. MSCs were plated onto 24-well plates in 500 μl of DMEM medium on day 0 and transfected with TRAIL-bearing or control plasmid on day1. On day 2, the cultures were washed twice with PBS and fixed with 4% paraformaldehyde for 10 min, and blocked with 2% bovine serum albumin at room temperature for 60 min. The cells were incubated with rabbit anti-hum TRAIL primary antibody (1:100) (Santa Cruz Biotechnology, Santa Cruz, CA, USA). After rinsing with PBS three times, cells were incubated with secondary antibodies Cy3-conjugated mouse anti-rabbit IgG 1:200 (Santa Cruz Biotechnology) for 60 min. Cell nucleoli was stained with DAPI. The images were taken using a fluorescence microscope (Leica DMIRE2).

TRAIL expression in MSCs. MSCs were plated in 6-well plates on day 0, transfected on day 1 and harvested on day 2. Total RNA and proteins were isolated for RT-PCR (A) and immunoblotting (B) analysis, respectively. 1: TRAIL transfection; 2: control plasmid transfection; 3: control.
Immunoblotting analysis. Immunoblotting analysis was used to detect the cellular expression of TRAIL in the MSCs. As previously described (13), MSCs transfeted with TRAIL or mock-infected were harvested in lysis solution. Whole-cell lysates (50 μg) were separated through 12% denaturing SDS-polyacrylamide gel electrophoresis and transferred to a nitrocellulose membrane. The membrane was incubated overnight with rabbit anti-TRAIL antibody or rabbit anti-β-actin antibody (1:1000), (Santa Cruz Biotechnology). This was followed by a 1-h incubation with goat-anti-rabbit IgG conjugated to horseradish peroxidase (HRP) (1:2,500). The blot was developed using enhanced chemiluminescence detection (Boster Bio-engineering, Wuhan, China).
Flow cytometric analysis. The expression of TRAIL receptors on the C6 cells was detected by flow cytometry. Sub-confluent C6 cells were detached with 0.25% trypsin and washed with PBS. A total of 1-5×105 cells were resuspended in 200 μl PBS for each reaction, and then 10 μl of primary antibody solution (25 μg/ml) (antibodies to human DR4, DR5, DcR1, DcR2 or isotype IgG, R&D Systems, Minneapolis, MN, USA) was added to each eppendorf tube and incubated at 4°C for 30 min. The cells were washed 3 times with PBS and re-suspended in 200 μl PBS. Then, the phycoerythrin-conjugated secondary antibody (R&D Systems) was added to each reaction. The cells were incubated for 30 min at 4°C in the dark. After being washed 3 times with PBS, the cells were re-suspended with 500 μl PBS in FACS analysis tubes for flow cytometric analysis (FACSCalibur, Becton Dickinson, Heidelberg, Germany). The mean fluorescence intensity of each receptor was assessed on the live cell population. The reference gating location was determined by both isotype IgG and secondary antibody controls with the aid of CellQuest software.
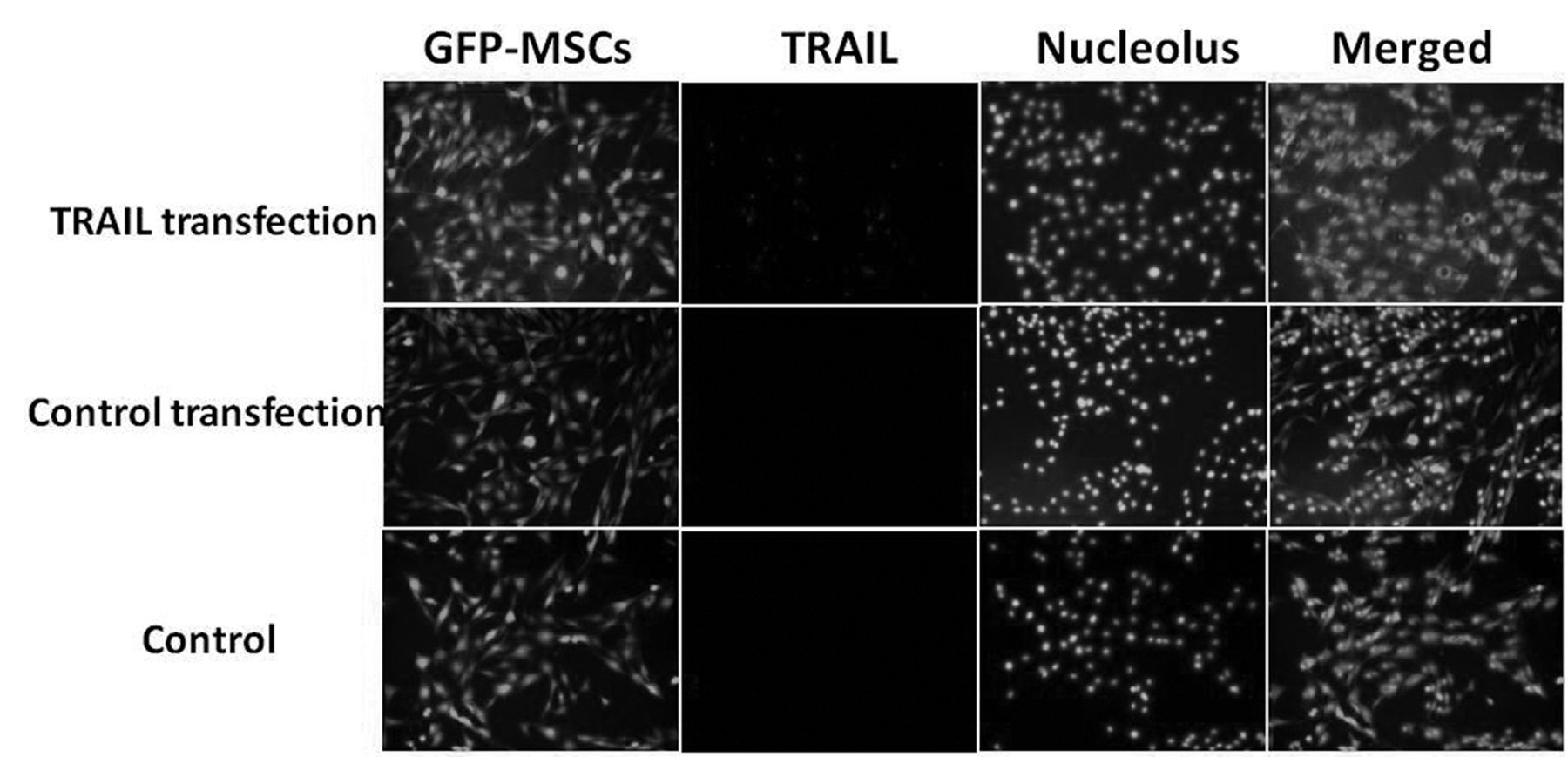
Immunofluorescent detection of TRAIL in MSCs. MSCs were plated in 12-well plates on day 0, transfected on day 1 and assessed by immunofluorescence on day 2. Original magnification, ×200.

FACS analysis of TRAIL receptors on C6 cells. The reference gate was set according to population and isotype IgG controls. The graph represents one of three parallel assessments.

Cell viability determined by MTT. A: The effects of TRAIL-transfection on MSCs' viability. MSCs were plated in 12-well plates on day 0 and transfected on day 1. MTT was performed on day 2. Data are presented as mean±SE (n=6). The comparison among groups showed no significant difference (p>0.05). B: The effects of TRAIL-engineered MSCs on C6 cell viability. MSCs were plated in 6-well plates on day 0 and transfected on day 1. On day 2, MSCs were harvested and co-cultured with C6 cells in 12-well plates. MTT was performed on day 3. Data are presented as mean±SE (n=6). *p<0.05 vs. control; 1, control; 2, control plasmid transfection; 3, TRAIL transfection.
Cytotoxicity assay. Cytotoxicities of TRAIL-transfected MSCs and C6 cells co-cultured with MSCs (40:60 ratio) were assessed with methylthiazol tetrazolium (MTT, Abcam, Toronto, ON, Canada) assay as per the manufacturer's instruction. The inhibition rate (IR) was defined as: IR (%)=(1-Sample A570/Control A570) ×100%. C6 cell viability was also detected with Hochest33258 staining followed by fluorescent microscopy analysis.
Statistical analysis. Numerical data were expressed as mean±standard error. Statistical differences between the means for the different groups were evaluated with SPSS13.0 (SPSS Inc., Chicago, IL, USA) using the Student's t-test with the level of significance at p<0.05.
Results
TRAIL expression in engineered MSCs. Figure 1A and 1B demonstrate the results of RT-PCR and immunoblotting of TRAIL expression in MSCs under different transfection conditions. TRAIL expression was verified at the mRNA and protein levels on TRAIL-transfected MSCs. The specific TRAIL expression in engineered MSCs was also confirmed by immunofluorescent detection (Figure 2).

The cytotoxic effects of TRAIL-engineered MSCs on C6 cells. The cell culture was as described in Figure 3B. Hochest33258 (10 μg/ml) was used to examine C6 apoptosis. A, Control; B, Control plasmid transfection; C, TRAIL trasfection. Original magnification, ×200.
TRAIL receptor expression in C6 cells. As shown in Figure 3, almost all tested C6 cells expressed the DR5 receptor and a small percentage of C6 cells had the DcR1 and DcR2 receptors.
The effects of TRAIL-transfection on MSCs' viability. As shown in Figure 4a, there was no significant effect of TRAIL transfection on MSCs' viability. Compared to non-transfected cells, IRs were 1.5% and 14.2% in plasmid-transfected and TRAIL-transfected cells, respectively (p>0.05).
The effects of TRAIL-engineered MSCs on C6 cell viability. The C6 cell viability was inhibited by MSCTRAIL, which is significantly different from control C6 cells and cells treated with control plasmid (p<0.05, Figure 4B). The inhibition rates (IRs) were 16.7%±0.11and 63.7%±0.12, respectively. The effects of MSCTRAIL on C6 cells were also identified with Hochest33258 staining. Representative images are displayed in Figure 5.
Discussion
Malignant glioma is one of the most refractory cancers, mainly due to the particularity of its location as well as its biological characteristics, such as (i) infiltrative nature, (ii) resistance to apoptosis, (iii) propensity for recurrence, and (iv) resistance to conventional therapies (2). MSC-mediated anticancer strategy holds great potential as a multiple-targeted and personalized therapy for glioblastomas. Glioblastoma-oriented migration of MSCs has been widely investigated both in vitro and in vivo (14-19). To make use of MSCs as anticancer agent vehicles, MSCs must be appropriately transfected with selected anticancer genes, and it is necessary to ensure that the viability of MSCs is not harmed by anticancer gene transfection. In the present study, MSCs retained their viability after transfection with control plasmids or TRAIL-bearing plasmids. This is in line with our previous liver cancer and pancreatic cancer studies (13, 20).
TRAIL and its receptors are important components of the extrinsic pathway of apoptosis. The presence of death receptors in cancer cells is a prerequisite determinant for TRAIL to induce apoptosis in targeted cancer cells (21). As shown in Figure 3, C6 cells were verified as DR5-dominant death receptors. A death domain in the intracellular region of DR4 and DR5 can recruit death-inducing signaling complex (DISC) upon TRAIL stimulation, and therefore, activate the downstream caspase cascade leading to cell death by apoptosis (12). The co-culture studies (Figures 4 and 5) exhibited the effects of TRAIL-engineered MSCs on C6 glioma cells by the MTT cytotoxicity assay and apoptotic staining. This proof-of-concept study provided direct evidence for TRAIL-engineered MSCs' cytotoxic effects on this particular cell type under direct co-culture conditions. However, it is worth noting that the sensitivity of tumor cells to TRAIL is not particularly consistent even under apparently identical conditions (22, 23). To render any MSC-mediated strategy clinically-meaningful, multiple anticancer agents should be included in the system.
In conclusion, mesenchymal stem cells were well-tolerant to the transfection of TRAIL-bearing vectors. The MSC-mediated cytotoxic effects of C6 cells are induced by TRAIL expression. The present study provided additional information contributing to the development of MSC-based strategies to treat patients with malignant gliomas.
Acknowledgements
This work was supported by the Taihe Hospital foundation.
Footnotes
-
↵* These authors contributed equally to this work.
- Received December 28, 2013.
- Revision received January 15, 2014.
- Accepted January 16, 2014.
- Copyright© 2014 International Institute of Anticancer Research (Dr. John G. Delinassios), All rights reserved





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}