


Abstract
Background: Adult T-cell leukemia/lymphoma (ATL) is an aggressive malignancy of peripheral T-lymphocytes and its prognosis still remains very poor. Materials and Methods: The potential of combining the Bcl-2 homology 3 mimetic ABT-737, which blocks Bcl-2, Bcl-XL, and Bcl-w, with either the proteasome inhibitor bortezomib or histone deacetylase (HDAC) inhibitor suberoylanilide hydroxamic acid (SAHA) to inhibit the growth of human T-lymphotropic virus type-I (HTLV-1) infected T-cell lines and its mechanism was further evaluated. Results: ABT-737 synergistically induced apoptosis when combined with either bortezomib or SAHA in HTLV-1 infected T-cell lines and fresh ATL cells. Bortezomib increased the expression of Noxa, which subsequently enhanced the formation of Mcl-1-Noxa complexes, resulting in the functional neutralization of Mcl-1, an inducer of resistance to ABT-737. On the other hand, SAHA reduced the expression of survivin, an anti-apoptotic molecule that confers drug resistance on ATL cells. Conclusion: The combination of ABT-737 with bortezomib or SAHA is promising for the treatment of ATL.
Adult T-cell leukemia/lymphoma (ATL) is an aggressive malignancy of peripheral T-lymphocytes associated with human T-cell lymphotropic virus type I (HTLV-1) (1, 2). The clinical subtypes of ATL have been divided into acute, lymphoma, chronic and smoldering (1), and a recent retrospective study showed that the median survival times of patients with these subtypes were 8.3, 10.6, 30.2 and 36.7 months and overall survival rates at 4 years were 11.4%, 16.2%, 35.0% and 43.2%, respectively (3). Novel therapeutic options such as allogeneic stem cell transplantation and anti-CCR4 monoclonal antibody have been recently introduced; however, the outcome of patients with ATL is still very poor and novel therapeutic approaches are urgently required.
Members of the Bcl-2 family proteins are critical regulators of apoptosis and interactions between anti-apoptotic and pro-apoptotic members are the major determinants of cell death and survival. ABT-737 is a small molecule Bcl-2 homology 3 (BH3) mimetic that binds to surface hydrophobic grooves of anti-apoptotic Bcl-2 family members and has been shown to strongly and selectively inhibit Bcl-2, Bcl-XL and Bcl-w, but not Mcl-1 or A1. In preclinical studies, ABT-737 exhibited single-agent activity and also increased the sensitivities of malignant lymphoma and small cell lung carcinoma cell lines to chemotherapeutics (4, 5). ABT-263 (Navitoclax), an analogue of ABT-737, was also reported to be effective on various cancer-derived cell lines in vitro, in vivo animal model, and in several clinical trials (6-11). We previously demonstrated that Bcl-2 and Bcl-XL, as well as the Mcl-1 protein, were highly expressed in ATL cells and HTLV-1 infected T-cell lines. Fresh tumor cells derived from ATL patients were less sensitive to ABT-737 than those derived from chronic lymphocytic leukemia (CLL) patients in vitro, however, ABT-737 synergistically enhanced apoptosis induced by current key conventional chemotherapeutics in HTLV-1 infected T-cell lines. Furthermore, the inhibition of Mcl-1 expression significantly enhanced the induction of apoptosis by ABT-737 (12). In addition, the nuclear factor-kappa-light-chain-enhancer of activated B (NF-κB) pathway, which induces anti-apoptotic and survival signals, was also shown to be constitutively activated in ATL cells (13).
The clinical activity of proteasome inhibitors against multiple myeloma has already been established. Proteasome inhibition is known to affect various intracellular signaling pathways, including NF-κB, cell-cycle regulation, modulation of Bcl-2 family members and accumulation of p53 (14). Previous studies have demonstrated that the proteasome inhibitor bortezomib induced cell death in HTLV-I infected T-cell lines and ATL cells, and its potential activity has also been reported in a xenograft murine model (15-18). We are currently conducting a multi-center phase II clinical trial of bortezomib in relapsed and refractory ATL patients (clinical trial registry number; UMIN000004061).
Histone deacetylase inhibitors (HDACi) are novel agents that inhibit the enzymatic activity of HDAC. HDACi can induce the acetylation of histone and non-histone proteins, which have been shown to affect various physiological and pathological processes controlling apoptosis, autophagy, cell cycle, immune response, inflammation and metabolism through its downstream molecular targets (19). HDACi were shown to influence the expression and activity of apoptotic proteins favoring a pro-apoptotic response and lowered the cellular apoptotic threshold (20). HDACi, such as suberoylanilide hydroxamic acid (SAHA), romidepsin and panobinostat, have shown promise in pre-clinical and/or clinical studies against T-cell malignancies including ATL (21, 22). However, the overexpression of anti-apoptotic Bcl-2 or Bcl-XL has been previously shown to reduce the efficacy of HDACi (23, 24). ABT-737 has been reported to sensitize Bcl-2 overexpressing tumors to HDACi-mediated apoptosis (25). Therefore, we postulated that ABT-737, combined with SAHA, may synergistically induce apoptosis in HTLV-1-infected T-cells.
In the present study, we examined the synergistic effects of the combination of ABT-737 with bortezomib or SAHA and further clarified the molecular sequences of HTLV-1 infected T-cell lines.
Materials and Methods
Cells. Two HTLV-1 infected T-cell lines MT-1 and MT-2 (kindly provided by Dr. Miyoshi I. (Kochi University, Nangoku, Japan)) were used in this study. MT-1 cells were established from the peripheral blood (PB) tumor cells of ATL patients (26), while MT-2 cells were established from cord blood T-cells by the co-cultivation of normal human cord lymphocytes and PB tumor cells of an ATL patient (27). Fresh PB ATL cells were separated by density sedimentation using LymphoPrep (PROGEN Biotechnik, Heidelberg Germany) from heparinized PB obtained from two acute-type ATL patients whose number of ATL cells comprised more than 85% of mononuclear cells, after obtaining informed consent. Cells were cultured at 37°C in Roswell Park Memorial Institute (RPMI) 1640 containing 15% fetal bovine serum (Sigma, St Louis, MO, USA), 2 μM L-glutamine, 100 U/ml penicillin and 100 μg/ml streptomycin (Gibco, Grand Island, NY, USA).
Reagents. An inhibitor of Bcl-2 family proteins, ABT-737 was provided by Abbott Laboratories (Abbott Park, IL, USA). The proteasome inhibitor bortezomib, HDACi SAHA and survivin inhibitor YM-155 were obtained from Toronto Research Chemicals Inc. (Ontario, Canada), Cayman Chemical (Michigan, IL, USA) and Selleck Chemicals (Houston, TX, USA), respectively. The pan-caspase inhibitor z-VAD-fmk (Z-VAD) was obtained from Bachem (Bubendorf, Switzerland). The Cell Counting Kit-8 (Dojindo, Kumamoto, Japan) was used to assess cellular proliferation by a colorimetric assay.
Detection of apoptosis. APO 2.7 staining (Immunotech, Marseille, France) was used to determine apoptosis and was evaluated using an EPICS XL flow cytometer (Beckman Coulter, Hialeah, FL, USA).
Western blotting. Western blotting was performed as previously described (28) with the following antibodies: anti-caspase 3, -caspase 9, -PARP, -Bcl-2, -Bcl-XL, -Bcl-w and -survivin (Cell Signaling, Danvers, MA, USA), and -Noxa and -Mcl-1 (Santa Cruz Biotechnology city, CA, USA). Immunoblotting with anti-α-Tubulin (Cell Signaling) confirmed equivalent protein loading.
Immunoprecipitation. Cells cultured under the indicated conditions were harvested, washed twice with ice-cold PBS, lysed in radioimmunoprecipitation assay (RIPA) buffer (Wako Pure Chemical Industries, Osaka, Japan), incubated with 2 μg anti-Mcl-1 antibody and 15 μl protein G-Sepharose was added. The immunocomplexes were collected and detected by Western blotting.
Analysis of drug synergy. The effects of combining ABT-737 with either bortezomib or SAHA were evaluated using the Chou-Talalay method to determine the combination index using the CalcuSyn software (Biosoft, Ferguson, MO, USA). Each affected fraction (Fa) was calculated by comparing the absorbance values of drug-treated wells measured by a colorimetric assay, to the absorbance of control wells. The drug concentration that induced Fa=0.25 signified a 75% decrease in absorbance and growth (i.e., IC25 concentration). Background absorbance was set at Fa=1. Based on this approach, combination index (CI) values of <0.9 were considered synergistic, >1.1 were antagonistic and values of 0.9 to 1.1 were additive (29, 30).
Results
Either bortezomib or SAHA synergistically enhanced the effects of ABT-737. We first examined the single-agent activity of ABT-737, bortezomib or SAHA on HTLV-1 infected T-cell lines by the colorimetric assay and showed that they inhibited the growth of MT-1 and MT-2 cells (Figure 1A). We then investigated the effects of combining ABT-737 with either bortezomib or SAHA. Dose-effect and Fa-CI plots revealing the effects of fixed drug ratio combinations are shown in Figure 1B. The combination of ABT-737 with bortezomib or SAHA displayed a strong synergism (CI<0.9) for inhibiting the growth of MT-1 and MT-2 cells.
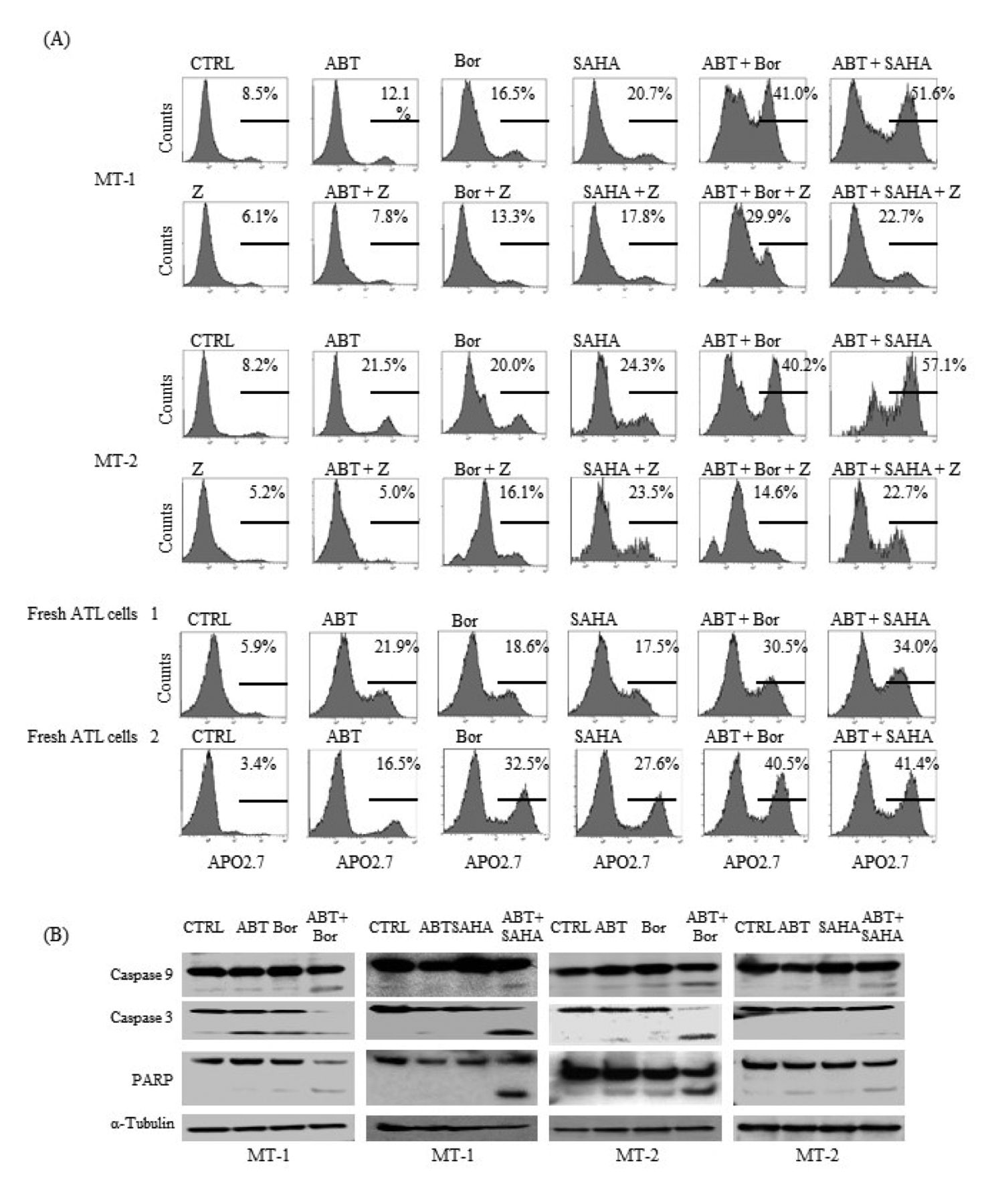
To clarify the in vitro anti-tumor effects achieved by these combinations, we examined the induction of apoptosis in MT-1, MT-2 cells and fresh ATL cells, treated by ABT-737 with or without bortezomib or SAHA. Apoptosis induced by ABT-737 was significantly enhanced by either bortezomib or SAHA. Of note, it was accompanied by the cleavage of caspase 3, caspase 9 and PARP, and was blocked by the pan-caspase inhibitor Z-VAD in MT-1 and MT-2 cells (Figure 2A and B).
Bortezomib induced the expression of Noxa. To explore the mechanism of the synergistic effects of ABT-737 and bortezomib, we examined the modulation of Bcl-2 family proteins in MT-1 and MT-2 cells. Bortezomib induced the expression of Noxa and Mcl-1 without modulating the expression of Bcl-2, Bcl-w or Bcl-XL (Figure 3A). Immunoprecipitation using the anti-Mcl-1 antibody revealed the enhanced formation of the Mcl-1-Noxa complex in both cell lines treated with bortezomib-alone and ABT-737 combined with bortezomib (Figure 3B).

Combination of ABT-737 with bortezomib or SAHA synergistically inhibited the growth of HTLV-1-infected T-cell lines. (A) The growth inhibition of MT-1 and MT-2 cells by ABT-737, bortezomib, and SAHA assessed by a colorimetric assay after a 72-h culture. Data represent means±SD (standard deviation) of 3 independent experiments. (B) MT-1 and MT-2 cells were treated with ABT-737 in combination with either bortezomib or SAHA for 72 h and the effects of the combined treatments were evaluated using the CalcuSyn software. Dose–effect and Fa-CI plots, illustrating the effects of fixed drug ratio combinations, are depicted. CI values <0.9 were considered synergistic, >1.1 were antagonistic and values of 0.9 to 1.1 were additive.
SAHA reduced the expression of survivin. We next elucidated the mechanism of synergy between ABT-737 and SAHA. No significant change was observed in the expression of Bcl-2, Bcl-w or Bcl-XL. SAH-alone and ABT-737 combined with SAHA slightly decreased the expression of Mcl-1. The expression of survivin was reduced by SAHA and was significantly reduced by combining SAHA with ABT-737 (Figure 4). To confirm the biological relevance of survivin in ABT-737-induced apoptosis, we demonstrated that the small-molecule inhibitor of survivin YM-155 significantly enhanced apoptosis induced by ABT-737 (Figure 5).

Combination of ABT-737 and bortezomib or SAHA synergistically induced apoptosis in HTLV-1-infected T-cell lines and fresh ATL cells. (A) MT-1 and MT-2 cells were treated by ABT-737 (1 μM), bortezomib (4 nM), SAHA (2 μM) or by their combination in the absence or presence of Z-VAD (25μM) for 72 h. Fresh ATL cells were treated by ABT-737 (1 μM), bortezomib (4 nM), SAHA (1 μM) or by their combination for 72 h. The induction of apoptosis was assessed by using the APO2.7 assay. The percentage of APO2.7-positive cells is shown. Data are representative of two independent experiments. CTRL, Control; ABT, ABT-737; Bor, bortezomib; Z, Z-VAD. (B) Cells were treated with ABT (1 μM), Bor (4 nM), SAHA (2 μM) or their combination for 24 h. Whole-cell lysates were subjected to western blotting to assess the cleavage of caspase 9, caspase 3 and PARP. CTRL, Control; ABT, ABT-737; Bor, bortezomib.

The effects of the combination treatment with ABT-737 and bortezomib on NOXA and Mcl-1 protein expression. (A) Cells were treated with ABT (1 μM), Bor (4 nM) or combination of both for 24 h. Whole-cell lysates were subjected to western blotting to assess Noxa, Mcl-1, Bcl-2, Bcl-w and Bcl-XL. CTRL, control; ABT, ABT-737; Bor, bortezomib. (B) Cells were treated with ABT (1 μM), Bor (4 nM) or combination of both for 24 h. Whole-cell lysates were immunoprecipitated with the anti-Mcl-1 antibody. Immunoprecipitates were then detected by western blotting using the anti-Noxa or anti-Mcl-1 antibody. CTRL, control; ABT, ABT-737; Bor, bortezomib.
Discussion
In the present study, we investigated the synergistic anti-ATL effects by combining ABT-737 and a proteasome inhibitor, bortezomib, or an HDACi, SAHA, in HTLV-1 infected T-cell lines and elucidated their mechanism of action.
ABT-737 in combination with bortezomib induced a synergistic apoptotic response in HTLV-1 infected T-cell lines and fresh ATL cells. Mcl-1 cleavage by bortezomib has been reported in multiple myeloma cells (31). The cleavage of Mcl-1 was not observed in our study; however, bortezomib up-regulated the expression of Noxa, resulting in the enhanced formation of Mcl-1-Noxa complexes. Because the BH3 domain of Noxa has been shown to antagonize Mcl-1, the up-regulation of Noxa resulted in the functional repression of Mcl-1 (32). Thus, the mechanism that enhances the activity of ABT-737 by bortezomib is suggested to cancel the anti-apoptotic effect of Mcl-1.

The effect of combination of ABT-737 and SAHA on survivin levels. Cells were treated with ABT (1 μM), SAHA (2 μM) or their combination for 24 h. Whole-cell lysates were subjected to Western blotting to assess survivin, Bcl-2, Bcl-w, Bcl-XL and Mcl-1. CTRL, control; ABT, ABT-737=Bor, bortezomib.

Inhibition of survivin sensitizes HTLV-1-infected T-cell lines to ABT-737. Cells were treated with ABT (1 μM), YM-155 (10 nM) or their combination for 72 h and induction of apoptosis was assessed using an APO2.7 assay. The percentage of APO2.7-positive cells is shown. CTRL, Control; ABT, ABT-737.
The combination of ABT-737 and SAHA also synergistically enhanced the induction of apoptosis in HTLV-1-infected T-cells by down-regulating the expression of survivin. Caspases are regulated by several intrinsic inhibitors of apoptosis proteins, in which survivin is a major one. Previous studies have shown that survivin induced resistance to ABT-737 (33, 34). In ATL, survivin has been suggested to be an important anti-apoptotic molecule that confers drug resistance, and high mRNA expression of survivin was shown to be a risk factor for the prognosis (35, 36). We also demonstrated that the inhibition of survivin by YM-155 sensitized MT-1 and MT-2 cells for induction of apoptosis by ABT-737. Our results suggest that SAHA increases sensitivity to ABT-737 by down-regulating the expression of survivin. The down-regulation of Mcl-1 was less significant than that of survivin, but may also contribute to the induction of apoptosis. Consequently, the combination of ABT-737 and SAHA is a promising treatment that targets multiple anti-apoptotic molecules such as Bcl-2, Bcl-XL, Bcl-w, Mcl-1 and survivin in HTLV-1 infected T-cells.
ABT-737 is a known inhibitor of some anti-apoptotic proteins. Bortezomib targets the proteasome/ubiquitin pathway, which is active on apoptosis pathway regulators, while SAHA targets histone and non-histone deacetylases and indirectly modulates apoptosis. Numerous regulators are known to be involved in apoptotic pathways; therefore, inhibiting a single pathway is not sufficient. ABT-737, bortezomib and SAHA by themselves exhibited limited activity against HTLV-1 infected T-cell lines. However, ABT-737 combined with bortezomib or SAHA synergistically enhanced the induction of apoptosis because they could compensate for the functional deficits of each other. Taken together, these novel strategies targeting multiple apoptotic pathways appear promising for ATL but should be validated by in vivo animal model and clinical studies.
Acknowledgements
This work was supported by Health and Labor Sciences Research Grants for Clinical Research (H23-rinkensui-ippan-011) from the Ministry of Health, Labour and Welfare of Japan, and funds (No.:127006) from the Central Research Institute of Fukuoka University (K.I.).
Footnotes
-
Conflicts of Interest
The Authors declare no conflicts of interest.
- Received July 2, 2014.
- Revision received July 22, 2014.
- Accepted July 24, 2014.
- Copyright© 2014 International Institute of Anticancer Research (Dr. John G. Delinassios), All rights reserved
References
In this issue





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Jump to section
Related Articles
Cited By...
- No citing articles found.