


Abstract
Background: Cytoplasmic PLA2 (cPLA2) has been shown to be the major enzyme responsible for arachidonic acid (AA) release. Because of this key role of cPLA2 in AA production, cPLA2 involvement in tumorigenesis has been suggested. However, contradictory data are found in the literature. Additionally, little is known regarding the role of cPLA2 in pulmonary tumor metastasis. Materials and Methods: Tumor metastases were detected by lung colonization and angiogenesis was assayed as growth of blood vessels from subcutaneous tissue into an implanted matrigel of basement membrane. The matrix metalloproteinases (MMP)-2, and MMP-9 were detected by PCR with sequence-specific primers. Results: In this study, the effects of inhibitors of cPLA2, 5-lipoxygenase (5-LO), and cyclooxygenase (COX)-2 on pulmonary metastasis formation by B16F10 melanoma cells were investigated. All of these inhibitors reduced B16F10 pulmonary metastasis formation in a dose-dependent manner. Importantly, cPLA2, and 5-LO, and COX-2 inhibitors reduced platelet-activating factor-induced angiogenesis in an in vivo mouse model employing Matrigel injected subcutaneously, and also reduced expression of MMP-2 and MMP-9 in the lungs. Conclusion: This study demonstrates that cPLA2 metabolites play critical roles in tumor metastasis via the promotion, at least in part, of angiogenesis and MMP expression.
- Tumor metastasis
- cytoplasmic phospholipase A2
- 5-lipoxygenase
- cyclooxygenase-2
- angiogenesis
- metalloproteinase
Metastasis is a major cause of death in cancer patients and is one of the major obstacles to successful treatment. The array of mediators involved in tumor metastasis remains to be precisely defined. An example is the activation of eicosanoids in many types of cancer (1). Eicosanoids, which are products of the cyclooxygenase (COX) and lipoxygenase (LO) pathways, contribute to cancer progression by promoting cell proliferation, motility, invasion, and angiogenesis (2-4).
COX is a key enzyme in prostaglandin (PG) synthesis. COX converts free arachidonic acid (AA) to PGH2, which is further converted into a variety of PGs by different PG synthases. Among the three COX isoforms that have been identified so far, COX-2 is directly involved in colorectal tumor development (5, 6). COX-2 overexpression is observed in most colorectal adenocarcinomas and adenomas (7, 8) and promotes tumor development mainly via the generation of PGs, particularly PGE2 (9, 10).
The AA-transforming enzyme 5-LO catalyzes the conversion of AA into 5(S)-hydroperoxyeicosatetraenoic acid (5-HPETE) and leukotriene (LT) A4 (11). Accumulating evidence suggests a role for the 5-LO pathway in tumor cell proliferation and survival; 5-LO protein has been detected in cancer cell lines of animal and human origin (12-14).
Eicosanoids are synthesized from intracellular AA, which is released from membrane phospholipids by the action of phospholipase A2 (PLA2) (15, 16). Several types of PLA2 are expressed in human cells. Among them, the 85 kDa group IVA cytoplasmic PLA2 (cPLA2) has been shown to be the major enzyme responsible for AA release, and represents the rate-limiting step in eicosanoid production (17).
Because of this key role of cPLA2 in AA production, cPLA2 involvement in tumorigenesis has been suggested. However, contradictory data are found in the literature. cPLA2 was shown to be overexpressed in human intestinal tumors (18-20), and the knockout of cPLA2 gene dramatically reduced lung tumorigenesis in mice (21). By contrast, cPLA2 expression appears to be reduced in mouse colonic tumors (22), and the knockout of cPLA2 gene enhanced mouse colonic tumor development (23). Additionally, little is known regarding the role of cPLA2 in pulmonary tumor metastasis.
In this study, we have focused on the role of cPLA2 and its metabolites, especially metabolites formed by the action of 5-LO and COX-2, in a mouse model of pulmonary metastasis using a melanoma cell line.
Materials and Methods
Animals. Pathogen-free female C57BL/6 mice were purchased from Samtaco Inc. (Osan, Republic of Korea), and kept in our animal facility for at least 1 week before use. All mice were used at 7 to 8 weeks of age at the start of each experiment. All experimental animals used in this study were utilized under the protocol approved by the Institutional Animal Care and Use Committee of the Chonbuk National University Medical School.
Reagents. The cPLA2 inhibitor, arachidonyl trifluoromethyl ketone (AACOCF3), and the COX-2 inhibitor, NS-398, were purchased from Cayman Chemical (Ann Arbor, MI, USA). The 5-LO activating protein inhibitor, MK886, and 5-LO inhibitor, AA861, were purchased from Biomol Research Laboratories Inc. (Plymouth Meeting, PA, USA). Platelet-activating factor (PAF) (1-O-alkyl-2-acetyl-sn-glyceryl-3-phosphoryl-choline) was purchased from Sigma-Aldrich Co. (St. Louis, MO, USA). Matrigel, an extract of murine basement membrane proteins, consisting predominantly of laminin, collagen IV, heparin sulfate proteoglycans, and nidogen/entactin, was purchased from BD Biosciences (San Jose, CA, USA).
Cell culture. The B16F10 mouse melanoma cell line, which is metastatic in the lungs of C57BL/6 mice, was originally supplied by the Tumor Repository of the National Cancer Institute (Bethesda, MD, USA), and maintained in RPMI-1640 (Life Technologies, Grand Island, NY, USA) supplemented with 10% fetal bovine serum (Cambrex Co., Walkersville, MD, USA) at 37°C in a 5% CO2 atmosphere.
Lung colonization assay. A single-cell suspension of B16F10 cells (>95% viability by trypan blue exclusion assay) in 100 μl of phosphate-buffered saline were injected i.v. into the mice. Different concentrations of inhibitors were administered as follows: AACOCF3, i.p. 30 min prior to B16F10 cell or PAF injection; NS-398 and MK886, i.p. 1 h prior to B16F10 cell or PAF injection; AA861, subcutaneously (s.c.) 1 h prior to B16F10 cell or PAF injection. Lungs were removed 14 days after B16F10 cell injection and fixed in Bouin's solution (Sigma Chemical Co., St. Louis, MO, USA). The number of colonies on the lung surface was counted under a dissecting microscope. Each group included 5-8 mice per experiment.
Angiogenesis assay. Angiogenesis was assayed as the growth of blood vessels from subcutaneous tissue into an implanted solid gel of basement membrane containing the test sample as described previously (24). Matrigel (10 mg/ml), in liquid form at 4°C, was mixed with 64 U/ml heparin plus the experimental substances or vehicle alone and injected (0.1 ml) into the dorsal subcutaneous tissue of mice. Matrigel rapidly forms a solid gel at body temperature, trapping the factors, which allows slow release and prolonged exposure to surrounding tissues. After 6 days, mice were killed and gels were recovered and assayed for angiogenesis. Briefly, the assay works by measuring the amount of hemoglobin in the vessels that have invaded the Matrigel using the Drabkin reagent kit 525 (Sigma Chemical Co.). Matrigels were reliquified by being placed at 4°C on ice with red cell lysing reagent (Sigma Chemical Co.) for 24 h. After brief centrifugation, 20 μl of supernatant were added to 100 μl of Drabkin's solution. The mixture was allowed to stand for 30 min at room temperature, and its absorbance was measured at 540 nm. The results were expressed as hemoglobin concentration (g/dl).
Real time RT-PCR. RNA was prepared from the lungs as described previously (25). Reverse transcription was performed using 1 μl of total RNA in 10 μl of reaction mixture (Promega, Madison, WI, USA) containing oligo (dT)15 and avian myeloblastosis virus reverse transcriptase. PCR was performed on the Rotor-Gene 3000 System (Corbett Research, Morklake, Australia) using the SYBR Green PCR Master Mix Reagent Kit (Qiagen, Valencia, CA, USA). Mouse-specific primers used were as follows: MMP-2 for real-time PCR, 5′-CTGGAATGCCATCCCTGATAA-3′ and 5′-CAAACTT CACGCTCTTGAGACTTT-3′; MMP-2 for the visualization of the results, 5′-CTCAGATCCGTGGTGAGATCT-3′ and 5′-CTTTGG TTCTCCAGCTTCAGG-3′; MMP-9 for real-time PCR, 5′-TCGTGGCTCTAAGCCTGACC-3′ and 5′-GACACATAGTGGGA GGTGCT-3′; MMP-9 for the visualization of the results, 5′-ATCCAGTTTGGTGTCGCGGAGC-3′ and 5′-GAAGGGGAAG ACGCACAGCT-3′; β-actin, 5′-CTGAAGTCACCCATTGAACA TGGC-3′ and 5′-CAGAGCAGTAATCT CCTTCTGCA-3′. The relative levels of mRNA were calculated using the standard curve generated from sequential cDNA dilutions. The mean cycle threshold (Ct) values from quadruplicate measurements were used to calculate the gene expression, with normalization to β-actin as an internal control. Calculations of the relative levels of gene expression were conducted using the complementary computer software (Corbett Research, Morklake, Australia) employing a standard curve. The results were expressed as fold increase over untreated mice. cDNA, amplified by PCR (Perkin Elmer System 2400; Norwalk, CT, USA), was visualized after staining with ethidium bromide.
Statistical analysis. Data are presented as the mean±SE. Statistical significance was determined by one-way ANOVA test (StatView; Abacus Concepts Inc., Berkeley, CA, USA). All experiments were conducted two or more times. Reproducible results were obtained and representative data are therefore shown in the figures.
Results
cPLA2 inhibitor inhibits experimental pulmonary metastasis of B16F10 cells. Different doses of the specific cPLA2 inhibitor, AACOCF3, were injected intraperitoneally (i.p.) 30 min before i.v. tumor cell injection (1.5×105 B16F10). Lungs were removed on day 14, and the number of surface colonies counted. The number of tumor colonies ranged from approximately 100 to 150. AACOCF3 inhibited tumor metastasis formation in a dose-dependent manner. It almost completely inhibited metastasis formation at doses of 10 and 20 mg/kg (Figure 1).

cPLA2 inhibitor inhibits experimental pulmonary metastasis of B16F10 cells. AACOCF3 was administered i.p. 30 min prior to B16F10 melanoma cell injection (1.5×105 cells/mouse, i.v.). Lungs were removed on day 14, and the number of surface colonies was determined. Each group includes 12 mice from 2 independent experiments. *P<0.05 and **P<0.01 compared with the control group. Values are expressed as the mean±SE.
5-LO and COX-2 inhibitors inhibit experimental pulmonary metastasis of B16F10 cells. The suppression of colonies shown in Figure 1 suggests that cPLA2 metabolites, such as LTs, PGs, and thromboxanes, the expressions of which are regulated by 5-LO and COX-2, are involved in this process. Therefore, the effects of 5-LO and COX-2 inhibitors on pulmonary metastasis of B16F10 cells were examined. Different doses of 5-LO and COX-2 inhibitors were injected i.p. 1 h before tumor cell injection. Inhibititors of COX-2, 5-LO activating factor, and 5-LO caused dose-dependent reduction of metastasis formation (Figure 2), indicating that metabolites formed by the action of COX-2 and 5-LO are involved in pulmonary metastasis of melanoma tumor cells.
Effects of cPLA2, 5-LO, and COX-2 inhibitors on angiogenesis and expression of MMP-2 and MMP-9. We have previously shown that PAF is a potent inducer of the transcription factor, nuclear factor (NF)-κB (26, 27). As a result of NF-κB activation, PAF is associated with the enhancement of angiogenesis (28), and MMP expression (25), in addition to cPLA2 activity (27, 29). Based on these findings, we examined the in vivo effects of cPLA2, 5-LO, and COX-2 on PAF-induced angiogenesis, in addition to MMP-2 and -9 expression.
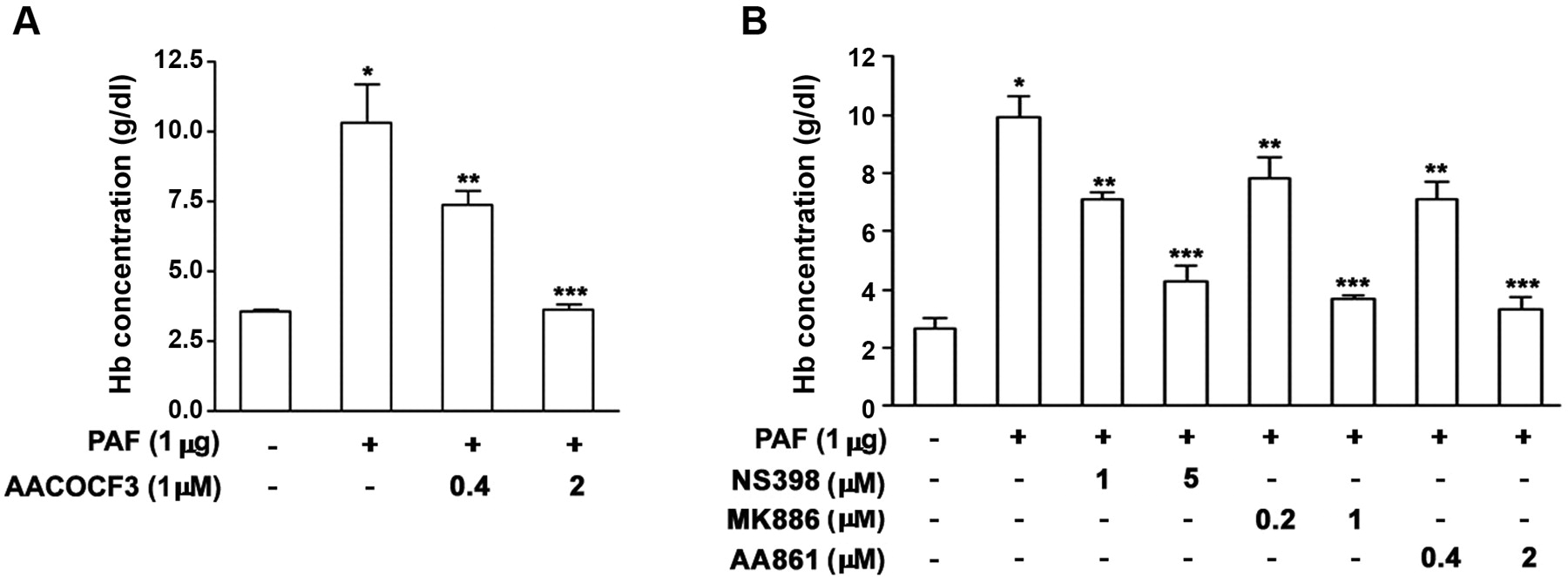
Matrigel containing PAF caused significant angiogenesis, which was inhibited by the inhibitors of cPLA2 (Figure 3A), COX-2, 5-LO activating factor, and 5-LO (Figure 3B).
PAF caused increases in the expression of MMP-2 and MMP-9 in the lungs. Inhibitors of cPLA2, COX-2, and 5-LO reduced PAF-induced MMP expression (Figure 4).
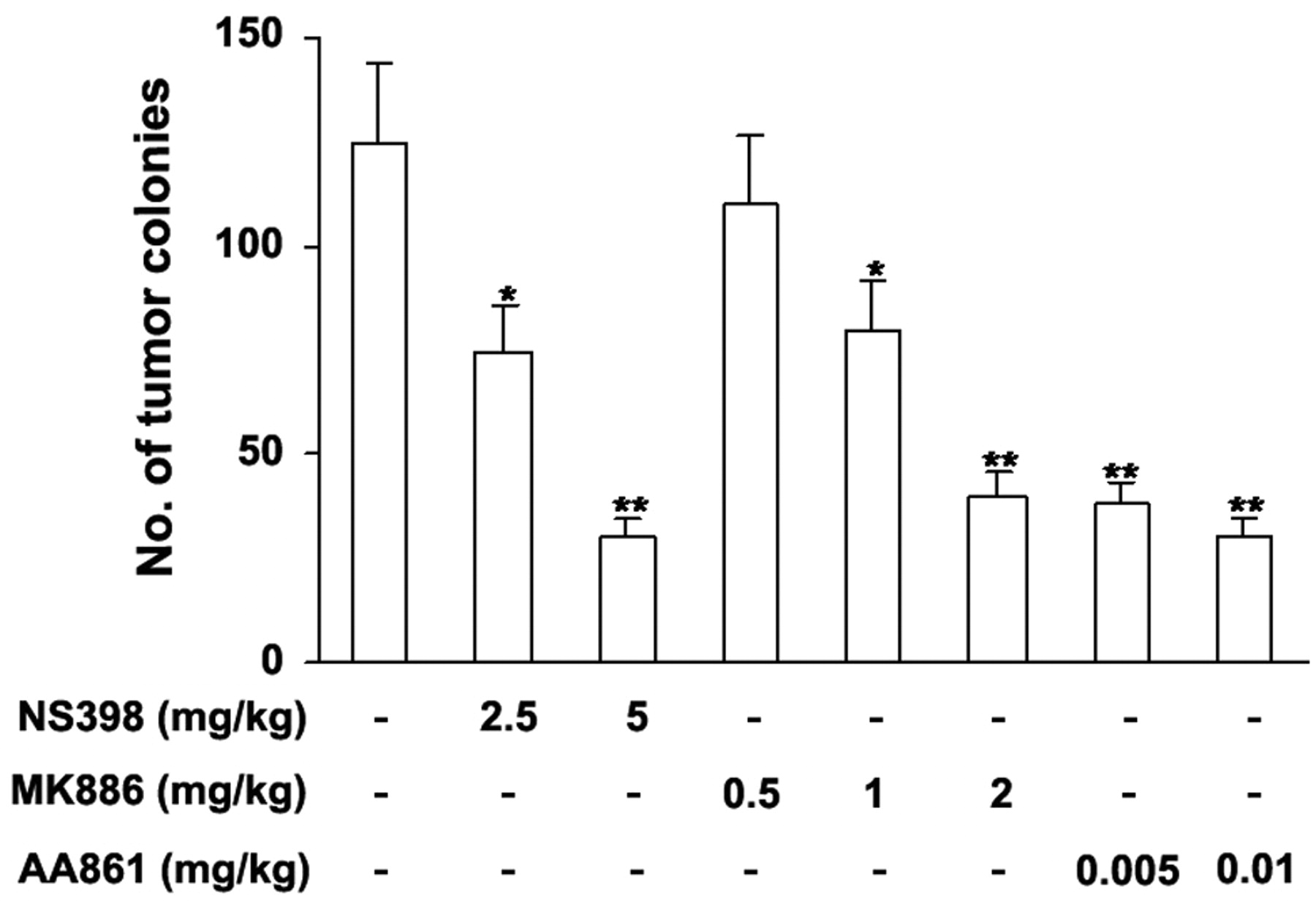
5-LO, and COX-2 inhibitors reduce experimental pulmonary metastasis of B16F10 cells. NS-398 was administered i.p. 1 h prior to B16F10 melanoma cell injection (1.5×105 cells/mouse, i.v.). MK886 and AA861 were administered i.p. and s.c., respectively, 1 h prior to B16F10 melanoma cell injection. Lungs were removed on day 14, and the number of surface colonies was determined. Each group includes 15 mice from 2 independent experiments. *P<0.05 and **P<0.01 compared with the control group. Values are expressed as the mean±SE.
Discussion
In this study, we focused on the roles of cPLA2 and its metabolites, especially those formed by the action of 5-LO and COX-2, in tumor metastasis. We found that the cPLA2 inhibitor exerted strong inhibitory activity against experimental pulmonary metastasis of B16F10 cells. Although studies showed that cPLA2 knockout mice exhibited dramatically reduced small bowel (30, 31) and lung tumorigenesis (32), little is known about the role of cPLA2 in tumor metastasis. Recently, in agreement with our data, it has been reported that metastasis to other lobes of the lung and to mediastinal lymph nodes, from primary tumor formed by mouse lung cancer cells injected directly into the lung, is decreased in cPLA2 knockout mice (33).
To investigate mechanisms for cPLA2-mediated metastasis, we examined the roles of COX-2 and 5-LO. We have demonstrated that 5-LO and COX-2 inhibitors reduced pulmonary metastasis of B16F10 cells, indicating that cPLA2 exerts its inhibition at least via 5-LO and COX-2. COX-2 overexpression is observed in most colorectal adeno-carcinomas and adenomas (34, 35), and promotes tumor development mainly by generation of PGs, particularly PGE 2 (36, 37). In addition, 5-LO inhibitor (38-40), and 5-LO activating protein (41, 42), along with LT antagonists (43, 44) essentially attenuate the effects attributed to 5-LO and its metabolites, thus blocking cell proliferation and inducing apoptosis in vitro and in vivo. However, the roles of COX-2 and 5-LO in tumor metastasis have not been well documented. In this regard, our data provide direct evidence that both COX-2 and 5-LO have a major role in tumor metastasis.

cPLA2, COX-2, and 5-LO inhibitors reduce PAF-induced angiogenesis. A: Matrigel plugs mixed with PAF (1 μg) and different concentrations of AACOCF3. B: NS398, MK886, or AA861 was mixed with the Matrigel and angiogenesis assays were performed on day 6. Results are expressed as hemoglobin concentration (g/dl). Each group includes 10 mice from 2 independent experiments *P<0.05 compared with the control group. **P<0.05 and ***P<0.01 compared with the PAF-treated group. Values are expressed as the mean±SE.

cPLA2, COX-2, and 5-LO inhibitors reduce PAF-induced expressions of MMP-2 and MMP-9 in the lungs. AACOCF3 was administered i.p. 30 min prior to PAF (1 μg/mouse) injection; NS-398, and MK886 were administered i.p. 1 h prior to PAF injection; AA861 was administered s.c. 1 h prior to PAF injection. The lungs were removed 4 h after PAF injection. Real-time RT-PCR was conducted as described in the Materials and Methods. A representative of three independent experiments is shown. *P<0.05 compared with the control group, **P<0.05 and ***P<0.01 compared with the PAF-treated group. Values are expressed as the mean±SE.
To determine the mechanisms for COX-2 and 5-LO inhibitor-mediated tumor metastasis, we investigated whether their inhibitors exert their effects by inhibiting angiogenesis and MMP expression. Neovascularization or angiogenesis is required to sustain primary tumor enlargement and metastasis growth. Induction of tumor angiogenesis is mediated by the increased production of various angiogenic molecules released by both tumor and host cells (45). This study demonstrated that COX-2 and 5-LO inhibitors significantly inhibited PAF-induced in vivo angiogenesis. Recent studies have indicated that neoangiogenesis, which is essential for tumor development, requires COX-2. COX-2 overexpressing colon cancer cells produce large amounts of proangiogenic factors, including vascular endothelial growth factor (VEGF) (46), a key regulator of endothelial cell migration and in vitro angiogenesis. Tumors implanted in COX-2 knockout mice display a reduction in vascular density and growth (47). In addition, 5-LO products, 5-HPETE and LTA4 but not LTB4, potently up-regulate VEGF transcription in a human malignant mesothelioma model (48).
The proteolytic degradation of extracellular matrix components is involved in both physiological and pathological processes, such as tissue remodeling, inflammation, tumor cell invasion, and tumor metastasis (49). Our study also demonstrated that 5-LO and COX-2 inhibitors reduced PAF-induced MMP-2 and MMP-9 expression. Given these observations, the fact that COX-2 and 5-LO inhibitors inhibited PAF-induced MMP-2 and MMP-9 expression suggests that promotion of extracellular matrix degradation by COX-2 and 5-LO products is likely associated with metastasis-augmenting activity. This hypothesis is further strengthened by the findings that a 5-LO activating protein inhibitor induced reduction in the gelatinolytic activity of MMP-2 (50), and 5-LO-deficient mice displayed decreased MMP-2 activity (51). In addition, it has been reported that COX-2 inhibitors inhibit the expression of MMP-2 and MMP-9 in prostate cancer (52) and MMP-2 levels were decreased in non-small cell lung cancer cell lines treated with a COX-2-specific inhibitor (53).
In summary, this study demonstrates that metabolites formed by COX-2 and 5-LO play critical roles in tumor metastasis via the promotion, at least in part, of angiogenesis and MMP expression, and suggests that COX-2 and 5-LO inhibitors have promising roles in the prevention of tumor metastasis.
Acknowledgements
This study was supported by a grant of Seoul Medical Center Research Institute, and by the Korea Research Foundation Grant funded by the Korean Government (KRF-313-C00872).
Footnotes
-
↵* These Authors contributed equally to this work.
- Received January 15, 2010.
- Revision received June 10, 2010.
- Accepted June 16, 2010.
- Copyright© 2010 International Institute of Anticancer Research (Dr. John G. Delinassios), All rights reserved





{kind=link}
{kind=link}
{kind=link}
{kind=link}