


Abstract
Aim: To investigate the expression and significance of glypican-3 (GPC3) in human hepatocellular carcinoma (HCC). Materials and Methods: DNA chips were used to measure the expression of mRNAs for members of the glypican and syndecan families of heparan sulfate proteoglycans (HSPGs) in normal liver tissue, non-tumor tissues and HCC. GPC3 protein expression was investigated by immunohistochemical staining in the tissues samples and Western blotting in human HCC cell lines. In addition, the levels of GPC3 protein in the blood were determined by ELISA. Results: Only the expression of GPC3 was found to be markedly elevated in HCCs. In the human HCC cell lines, GPC3 expression was consistently observed, and was mainly located in the cell membrane and cytoplasm. Immunohistochemistry showed a tendency for overall staining of the cytoplasm of cells in the liver carcinoma tissues, but the cell membrane was preferentially stained in poorly differentiated HCC when compared with well-differentiated HCC. Moreover, the cell membrane was preferentially stained in metastatic lesions of HCC when compared with primary HCC lesions. Non-tumor tissues and cholangiocellular carcinoma tissues were not stained. In addition, using HepG2 cells, AG490 and piceatannol, which are signal transducer and activator of transcription 3 (STAT3) inhibitors, each increased the amount of GPC3 mRNA expressed. Assay of the circulating levels of GPC3 protein in chronic liver disease and HCC found that serum GPC3 protein levels were significantly elevated in the latter. Conclusion: GPC3 is highly expressed in HCC, and its expression pattern differs according to the degree of cell differentiation. In addition, the expression of GPC3 is regulated by Janus kinase-STAT signaling. GPC3 shows potential as a tumor biomarker for HCC that can be used for molecularly targeted therapy.
Hepatocellular carcinoma (HCC) is one of the most common carcinomas worldwide. The prognosis of HCC is generally poor, with a low 5-year-survival rate, while the recurrence rate is high, and existing treatment modalities, whether drug- or surgery-based, have their limitations. It is thought that various clinicopathological factors, such as intrahepatic metastasis, a low degree of differentiation, large tumor size and portal vein invasion, worsen the prognosis of HCC patients. However, cases of advanced HCC are not unusual (1). Thus, it has become increasingly important to identify biomarkers that are specifically expressed in HCC.
Heparan sulfate proteoglycans (HSPGs) are present on the cell surface and in the extracellular matrix of all tissues. They are important structural components of the basal membrane, which forms the cellular exoskeleton and is the boundary membrane of tissues. Cell surface HSPGs function as receptors for cell adhesion and growth factors and are involved in the regulation of cell proliferation and differentiation, as well as cell motility. The basal membrane is enzymatically and/or physically disrupted by hematogenous metastatic cancer cells that invade blood vessels from a primary lesion and then extravasate from the blood into the extravascular tissues, and inflammatory cells that exit from blood vessels and infiltrate the surrounding tissues. The liberation of angiogenic and/or growth factors that are capable of binding to heparan sulfate then leads to the induction of angiogenesis, which in turn promotes further infiltration and proliferation of cancer cells themselves. It is thought that this sequence of events culminates in blood-borne metastasis. Accordingly, HSPGs are thought to play key roles in the proliferation and progression of carcinomas (2-3).
Glypican-3 (GPC3) is one of six of the glypican family of HSPGs and binds to cell membranes via a glycosylphosphatidylinositol (GPI) anchor that is present on cell membranes. The functions of GPC3 are thought to include important roles in cell growth, differentiation and motility. In other words, GPC3 plays a key role in relation to signaling with growth factors, thereby regulating the proliferative activity of cancer cells. Hsu et al. have investigated genes that show increased expression in HCC and reported that the GPC3 mRNA levels were significantly higher in HCC than in normal liver tissues and non-tumors liver tissues (4). In addition, at the protein level, the expression of GPC3 has been reported to be elevated in HCC (5-7). Moreover, ELISA studies have confirmed that GPC3 was not present in sera from healthy subjects or patients with hepatitis, but was significantly elevated in sera from 53% of patients with HCC (5).
On the other hand, GPC3 inhibits cell growth and induces apoptosis in some types of tumor cell. The expression of GPC3 has been reported to be down-regulated in tumors arising from various organs. In particular, GPC3 was reported to show inhibitory activity in breast cancer, ovarian cancer and malignant mesothelioma (8-11). However, numerous aspects remain to be clarified with regard to the expression and function of GPC3 in relation to the various types of cancer.
The present study was thus designed in consideration of the above background to investigate whether various members of the glypican and syndecan families of HSPGs are expressed in the normal liver, HCC-associated hepatitis and hepatitis without HCC (12, 13).
Materials and Methods
Cells and HCC tissues. The human HCC cell lines HepG2 (JCRB 1054), HLE (JCRB 0404) and Huh7 (JCRB 0403) were purchased from the Human Science Research Resource Bank (Osaka, Japan). The HCC cell line SK-Hep1 was purchased from the American Type Culture Collection (Rockville, MD, USA). All the cells were cultured in Dulbecco's modified Eagle's medium (Dainippon Pharmaceutical Co., Ltd., Osaka, Japan) at 37°C, supplemented with 1% penicillin/streptomycin (Gibco Brl, Grand Island, NY, USA) and 10% heat-inactivated fetal calf serum (Gibco Brl).
A total of 59 HCC tissues (43 tissues with primary HCC and 16 tissues with metastasis from HCC tissues), 9 corresponding non-tumor tissues and 4 tissues with cholangiocellular carcinoma were obtained from tissue array slides (SuperBioChips Laboratories, Seoul, Korea). Informed consent was obtained from all the patients for the subsequent use of resected tissues. The non-tumor liver tissues consisted of 4 cirrhotic specimens and 5 chronic hepatitis specimens. The resected tissues were frozen immediately at −80°C or fixed in 10% formalin.
Profiling analysis of GPC3 and syndecan recognized by viral components (DNA chips). The DNA chips were employed to analyze the expression of glypican-1-5 and syndecan-1, -3 and -4 in normal liver, HCC and non-tumor liver tissues. The total RNA was extracted using TRlzol (Life Technologies, Rockville, MD, USA). The RNA concentration and purity were determined using a NanoDrop ND-1000 spectrophotometer (NanoDrop, Rockland, DE, USA) and 2100 Bioanalyser (Agilent, Palo Alto, CA, USA), respectively. The synthesis of biotin-labeled cDNA probes, and the hybridization, washing, staining, and scanning steps were performed according to the manufacturer's instructions (Affymetrix Inc, Santa Clara, CA, USA).
Immunoblotting. The expression of GPC3 in the human HCC cell lines (HepG2, HLE and Huh7) was analyzed by immunoblotting. Briefly, after incubation on 6-well plates (Nunc™ Brand Products, NY, USA) for 48 h, the cells were washed twice with phosphate-buffered saline (PBS) and then lysed by the addition of SDS sample buffer (50 mmol/l Tris-HCl, pH 6.8, 2.5% SDS, 5% glycerol, 5% 2-mercaptoethanol and 0.01% bromophenol blue). Equal quantities of extracted proteins were separated by SDS-PAGE and were then transferred to poly vinylidene diflioride (PVDF) membranes (Millipore, Billerica, MA, USA). The blots were blocked by incubating in Tris-HCl (pH 7.5) containing 5% milk and 0.1% Tween 20 for 30 min at room temperature, and probed overnight at 4°C with primary antibodies. The following primary antibodies were used: anti-GPC3 monoclonal antibody (Imgenex, California, USA) and anti-α-tubulin monoclonal antibody (Oncogene Research Products, San Diego, CA, USA). The antibodies were diluted in 5% milk or in bovine serum albumin (BSA) in Tris-HCl (pH 7.5) containing 0.1% Tween 20. The immunoblots were then probed with horseradish peroxidase-conjugated anti-mouse immunoglobulin G (IgG) (diluted 1:1,000 in 1% milk or in BSA in Tris-HCl, pH 7.5) (Amersham Biosciences, Buckinghamshire, UK).
After the final wash, signals were detected using an ECL kit (Amersham Pharmacia Biotech).
In order to elucidate the subcellular localization of GPC3, cell lysates of the HepG2 cells were separated into cytosolic (F1), membrane/organelle (F2), nuclear (F3) and cytoskeletal matrix (F4) protein fractions using a ProteoExtract™ Subcellular Proteome Extraction Kit (Merck KGaA, Darmstadt, Germany) according to the manufacturer's instructions. Each fraction was examined for GPC3 expression by immunoblotting as described above.
Immunohistochemical staining. Immunohistochemical staining for GPC3 was performed on the HCC tissues and non-HCC tissues using the labeled streptavidin-biotin method (Dako Corp., Carpinteria, CA, USA). Deparaffinized sections were heated for 5 min at 120°C in a pressure cooker to reactivate the antigen, followed by treatment with 0.3% H2O2 in methanol for 20 min in order to abolish endogenous peroxidase activity. The sections were blocked with 1% BSA in PBS and covered with a 1:200 dilution of mouse anti-GPC3 monoclonal antibody in PBS (Santa Cruz Biotechnology, California, USA) overnight at 4°C. The sections were incubated with a second biotinylated antibody, followed by avidin-biotin-peroxidase complex (Vectastain ABC kit; Vector Laboratories, Burlingame, CA, USA) for 30 min. After washing, the sections were developed in a substrate solution of 0.01% 3,3′-diaminobenzidene-hydrogen peroxidase (DAB; Wako, Osaka, Japan) and counterstained for nuclei with 10% hematoxylin (Wako). The analysis of immunohistological staining was performed by independent researchers without the awareness of final diagnosis.
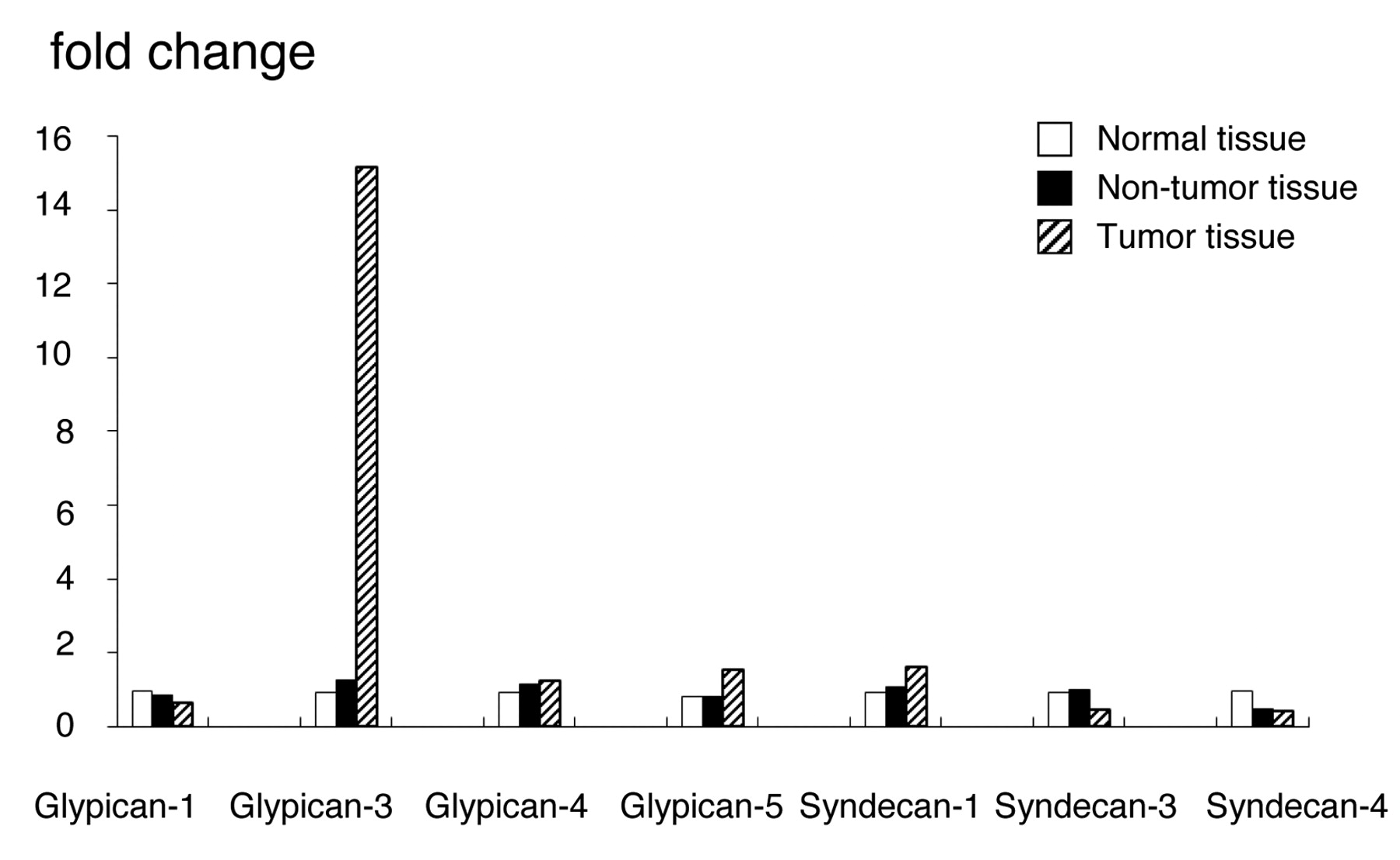
Gene expression of GPC and syndecans analyzed by DNA chip. Y axis indicates normalized expression values relative to normal tissues.
ELISA. The serum levels of GPC3 in patients with HCC and those with chronic liver disease were measured using a human GPC3 ELISA kit (BioMosaics Inc., Burlington, VT, USA) according to the manufacturer's instructions.
Jak-STAT inhibition experiment. In order to investigate the effects of STAT3 on the regulation of GPC3 expression, 100 μM AG490 (α-cyano-(3,4-dihydroxy)-N-benzylcinnamide) and piceatanol (3,4,3′,5′-tetrahydroxy-trans-stilbene) (Calbiochem, San Diego, CA, USA) which are inhibitors of STAT3, were added to HepG2 with standard cell culture conditions for 24 hours. GPC3 expression was then analyzed DNA chips
Statistical analysis. Unpaired Student's t-test was used to compare averages between groups, and chi-squared test and Fisher's exact probability test were used to compare independence.
Results
Expression of glypicans and syndecans in human liver. Among the glypicans, GPC3 was expressed at approximately 15-fold higher levels than glypican-1, -4 and -5. Moreover, its expression was similarly approximately 15-fold higher in HCC than in normal liver and non-tumor hepatic tissues. We could not obtain reliable results for glypican-2 expression. On the other hand, syndecans showed no significant differences in expression in any of the analyzed tissues (Figure 1). In addition, no changes in the level of GPC3 expression in normal liver tissue as a function of age was found over an age range of 20-59 years (data not shown).

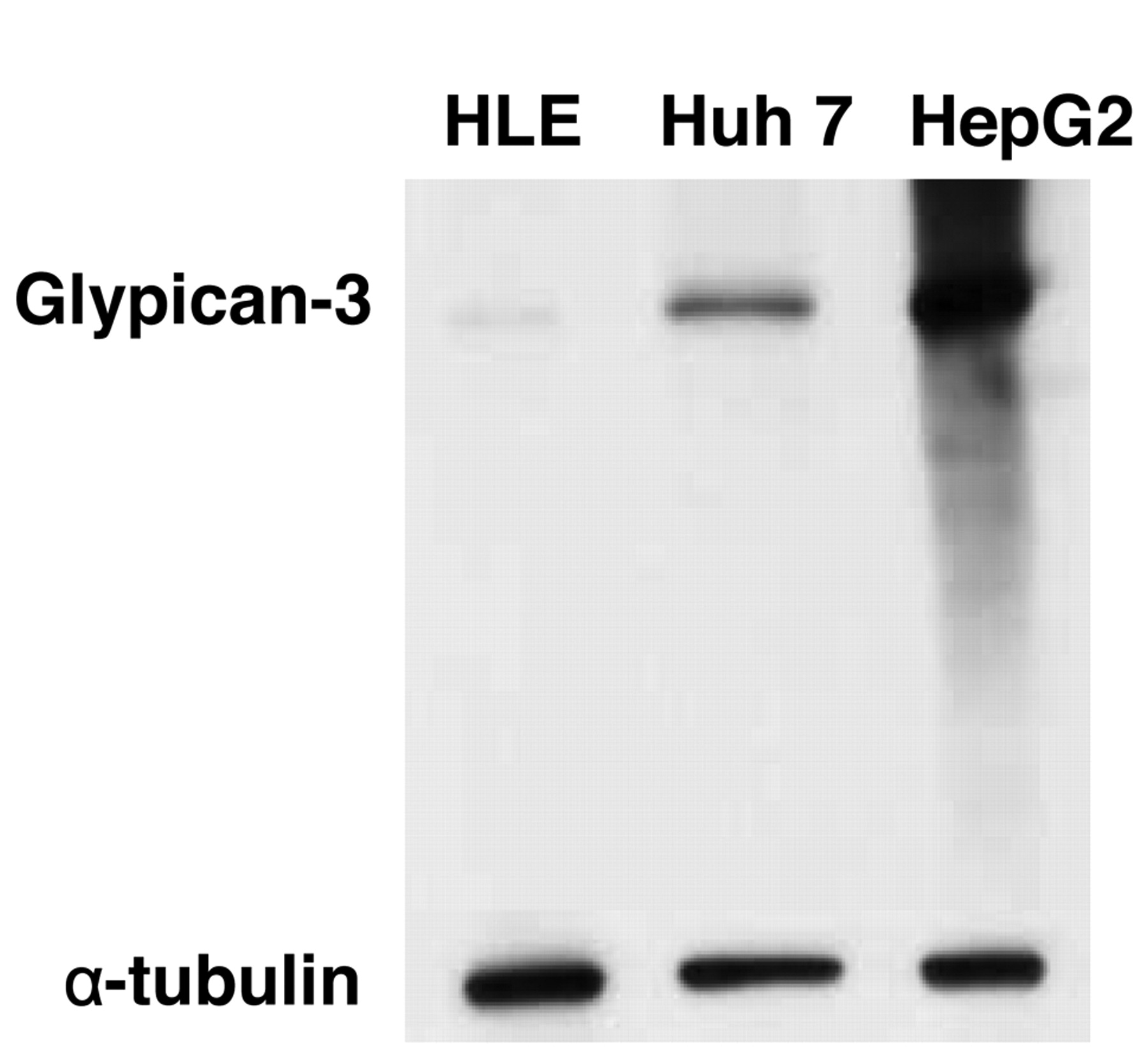
GPC3 expression in human HCC cell lines (HLE, Huh7 and HepG2), assessed by Western blot analysis.

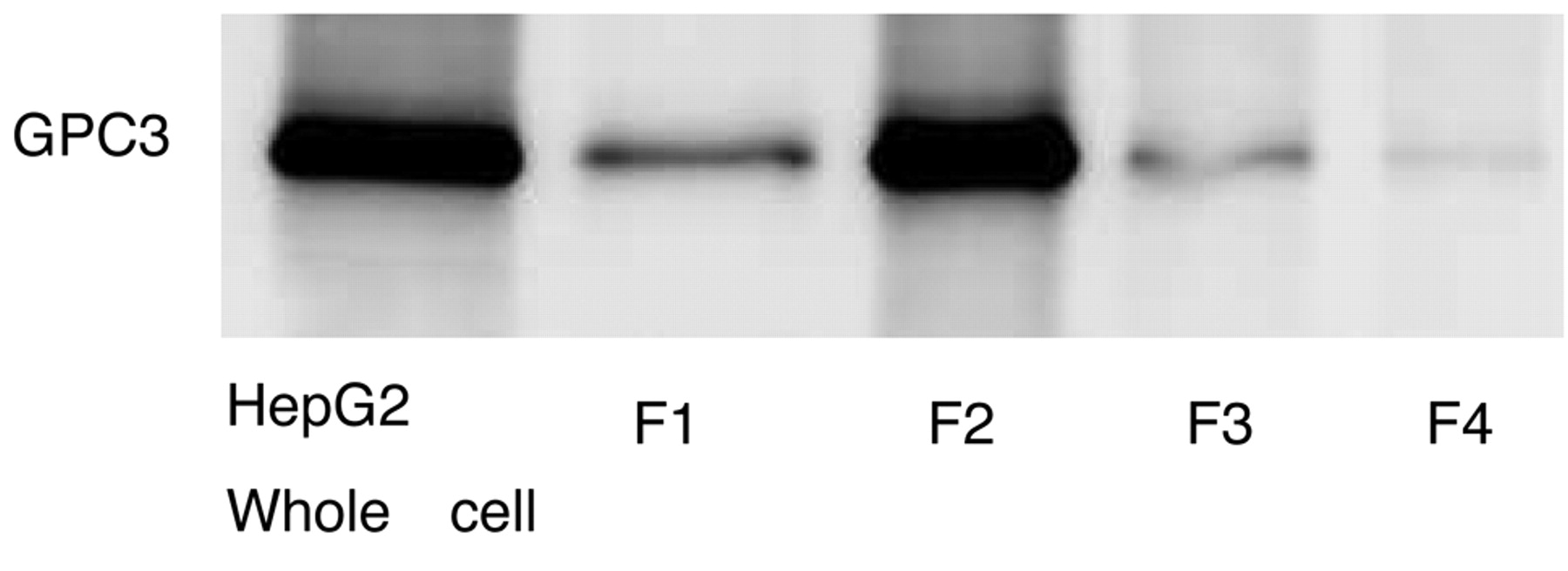
GPC3 expression analyzed by Western blot in HepG2 cells and F1: cytosolic, F2: membrane/organelle, F3: nuclear and F4: cytoskeletal matrix fractions.
Expression of GPC3 in HCC cells. GPC3 expression in several human HCC cell lines (HLE, Huh, HepG2) was confirmed by Western blotting (Figure 2). In addition, GPC3 was expressed in the cell membrane and cytoplasmic fractions of the HepG2 cells, with particularly strong expression in the cell membrane fraction (Figure 3).
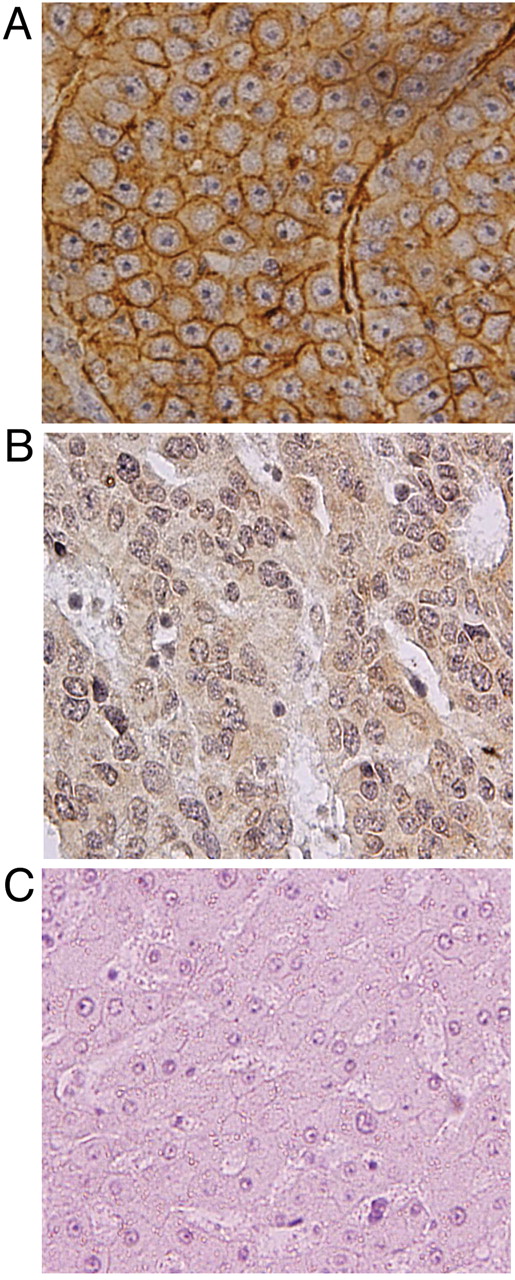
Expression of GPC3 in liver tissues. Almost no GPC3 expression was seen in the normal liver or non-tumor tissues, while strong positive staining was seen at the carcinoma sites. Figure 4A shows strong immunohistochemical staining of the cell membrane, while Figure 4B shows strong staining of the cytoplasm in HCC tissue samples. Figure 4C shows normal tissue, which was not stained.
The primary HCC tissues were classified into three groups by their degree of differentiation: well-differentiated HCC, moderately differentiated HCC and poorly differentiated HCC. The levels of GPC3 expression were analyzed in each group and in the metastatic lesions. The results are shown in Table I. The percentages of positive staining area were divided into three groups. In both the primary HCC and the metastatic lesions, the percentage of cases with 66% or greater positive staining for GPC3 was greater than the percentage with weak staining (~33%). In addition, the normal tissues were not stained at all.
The intensity of staining of the cell membrane and cytoplasm was divided into three levels and the results are shown in Table II. In both the metastatic sites and the primary HCC, there was a tendency for the cytoplasm to be readily stained throughout. The cell membrane was predominantly stained in poorly differentiated HCC when compared with well-differentiated HCC. Moreover, the cell membrane was predominantly stained in the metastatic lesions of HCC when compared with the primary HCC lesions. The cholangiocellular carcinoma tissues and normal tissues were not stained.
Serum levels of GPC3. Serum levels of GPC3 were significantly elevated (approximately 3-fold) in the patients with HCC compared to the patients with chronic liver disease (Figure 5).

A: Positive immunohistochemical stain for GPC3 with membranous pattern in HCC. B: Positive immunohistochemical stain for GPC3 with cytoplasmic pattern in HCC. C: Adjacent non-tumor tissue showing negative staining for GPC3.
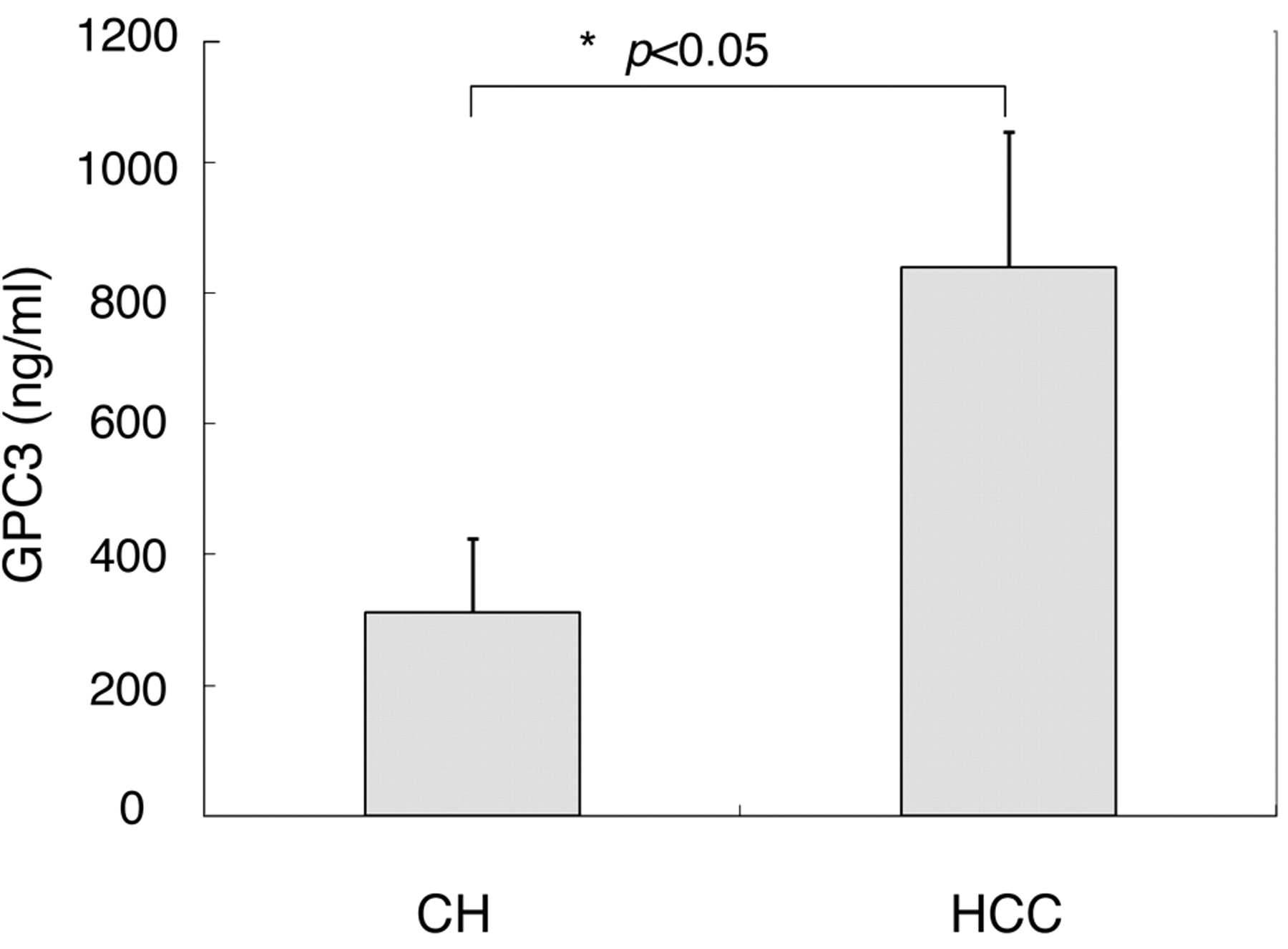
Serum GPC3 protein. CH: Chronic liver disease.
Regulation of GPC3 by Jak-STAT inhibitors. When the HepG2 cells were cultured with either AG490 or piceatanol the results showed that the level of GPC3 mRNA expression was higher than that in the controls (Figure 6).
Discussion
These present results indicated that glypicans, rather than syndecans, are involved in the onset and growth of HCC, with GPC3 playing a particularly large role.
In the present immunohistochemical staining study, all HCC specimens showed positivity for GPC3. However, nine normal tissues showed no positive staining for GPC3. Among the specimens that stained positively for GPC3, more showed 66.6% or greater stained area than showed 33.3% or less staining. These results indicated that GPC3 is expressed at a high rate and specifically in HCC. This was in agreement with the results of earlier similar studies, and GPC3 appears not to inhibit HCC, but to promote it (14). Liver cancer develops from tissues that normally express GPC3 only during the fetal stage, and it is thought that renewed GPC3 expression occurs in tandem with malignant transformation (6).
We found a tendency for greater GPC3 staining of the cytoplasm. Metastatic lesions also tended to show staining of the cytoplasm. Earlier reports of immunohistochemical staining of HCC documented a similar tendency for readily stained cytoplasm (14-16). It has been reported that for the expression of GPC3 in HCC, approximately half of the HCCs showed a mixture of cytoplasm and membrane staining, while the remaining half showed a cytoplasm-only pattern (1).
In the present study, the cell membrane was predominantly stained in poorly differentiated HCC when compared with well-differentiated HCC. The cytoplasmic staining did not show this tendency. Moreover, the cell membrane was predominantly stained in the metastatic HCC lesions when compared with the primary HCC lesions. Earlier reports had found no differences in GPC3 expression relative to the degree of HCC differentiation (17-18), while other report had found correlation between GPC3 expresson and the degree of HCC differentiation (19). However, very few investigations concerning the degree of HCC differentiation and differences in the localization of GPC3 have been reported.

The effects of AG 490, piceatanol, and GPC3 expression in Hep G2 cells.
GPC3 expression shown by extent of stained area.
In the present immunohistochemical staining study, with no exceptions, the cytoplasm was always stained in the specimens that stained positively for GPC3, regardless of the degree of differentiation and whether it was a metastatic or primary lesion. However, when the degree of GPC3 expression in each of the cytoplasmic, cell membrane and nuclear fractions, of the human HCC cell line was investigated there was strong expression in the cell membrane.
GPC3 expression shown by staining intensity in cytoplasm and membrane.
The reasons for this discrepancy in GPC3 staining are unclear. However, with regard to differentiation and metastasis, it is possible to conclude that there are differences in expression in the cell membrane.
GPC3 showed excellent sensitivity and specificity in the present experiments. However, GPC3 is secreted despite its being bound to the membrane via the GPI anchor, and it has been reported that it can be quantitatively detected in the serum by ELISA (6). Using the present sandwich ELISA technique, although the level of the GPC3 protein was significantly higher in the HCC patients, it was also present in small amounts in patients without cancer. This finding differed from the staining results. In addition, Nakatsura et al. reported that they were unable to detect GPC3 in the sera of patients with liver diseases other than HCC, healthy subjects or patients with other carcinomas (6). A more sensitive and specific assay method for GPC3 is thus necessary.
With regard to the GPC3 signal transduction pathway, it is said that glypican bound to the cell membrane modulates signals such as Wnts, Hedgehogs (Hedgehog as well as Wnts are one of the pathways in process of generation), fibroblast growth factors and bone morphogenetic proteins (20-25). However, on the other hand, there have been few reports concerning regulation of GPC3 expression.
STAT3 is involved in oncogenesis through the up-regulation of genes encoding apoptosis inhibitors (i.e., Bcl-xL, Mcl-1(Mcl-1 as well as Bcl-xL is a Bcl-2 family protein which can act as an apical molecule in apoptosis) and survivin), cell-cycle regulators (i.e., cyclin D1 and c-Myc(DNA binding protein concerning the regulation on metabolism of nucleic acid and the response to growth factor)), and inducers of angiogenesis (i.e., vascular endothelial growth factor). Therefore, STAT3 has recently drawn attention as a novel target for cancer therapy (26-28). Recent studies have suggested that in HCC, the Jak-STAT pathway is activated and inhibition of constitutive STAT3 activation may be a novel target for therapy (28-30). However, the role of Jak inhibitors in apoptosis and cell-cycle regulation is poorly understood in HCC. In the present study addition of either of the STAT3 inhibitors AG490 or piceatanol significantly increased the amount of GPC3 mRNA expressed in the cultures. These results indicated that poor control of the pathway mediated by STAT3 plays a role GPC3 expression.
In conclusion, GPC3 is specifically expressed in HCC, with strong immunohistochemical staining of the cell cytoplasm, whereas staining of the cell membrane is more evident in poorly differentiated specimens and specimens from metastatic lesions. Further study is needed with regard to the function of GPC3, but the findings to date indicate that this biomarker is a new molecular target for the treatment of HCC.
- Received August 9, 2010.
- Revision received October 18, 2010.
- Accepted October 19, 2010.
- Copyright© 2010 International Institute of Anticancer Research (Dr. John G. Delinassios), All rights reserved





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}