


Abstract
Tissue-specific expression of 25-hydroxyvitamin D-1α-hydroxylase (1α-OHase) and vitamin D-hydroxylase (24-OHase) may act as the pivotal link between 25-hydroxyvitamin D3 (25(OH)D3) serum levels and the anticancer effects of 1,25-dihydoxyvitamin D3 (1,25(OH)2D3) and alternative splicing of the enzymes may regulate their biological function. The expression of 24-OHase in cells and breast tissue was investigated and its splice variants were detected. The expression of 24-OHase RNA and protein was assessed by RT-PCR followed by Western blot. The expression of 24-OHase was reduced by about 57% in MCF-7 breast cancer cells, compared to MCF-10F benign breast cells. In the Western blot, a signal at 56 kDa was found and further bands were detected at 42 and 44 kDa. In the breast cancer tissue, the expression of 24-OHase was reduced by about 58% compared to benign tissue. However, in the Western blot, only one signal was found in the benign tissue at 56 kDa, while in malignant tissue, a further band was detected at 40 kDa. Alternative splicing of 24-OHase may lead to a catalytically dysfunctional enzyme and may lead to less reduction of the target protein.
Breast cancer is the most common and most life-threatening form of cancer among women in the U.S. and Europe (1). Many factors have been linked to various breast cancer risks including vitamin D3 synthesis in the skin due to sunlight exposure or dietary intake (2, 3). Several studies have shown that vitamin D3 might be inversely associated with the risk of breast cancer (4, 5). The biologically active metabolite of vitamin D3 is the secosteroid 1,25-dihydroxyvitamin D3 (1,25(OH)2D3). There are two principal enzymes involved in the formation of circulating 1,25(OH)2D3 from vitamin D3: the hepatic vitamin D-25-hydroxylase (25-OHase) and the renal 25-hydroxyvitamin D-1α-hydroxylase (1α-OHase, encoded by the cytochromeP27B1 gene) for vitamin D and 25-hydroxyvitamin D3 (25(OH)D3), respectively (6, 7). Both 25(OH)D3 and 1,25(OH)2D3 can be degraded through vitamin D 24-hydroxylase catalysis (24-OHase), encoded by the CYP24 gene (8).
In the kidney, 1α-OHase and 24-OHase form a classic feedback mechanism. Dietary intake and sunlight exposure significantly influence the circulating concentration of 25(OH)D3 which is considered to be a reliable indicator of vitamin D availability through the diet and synthesis in the skin (9). Conversely, the circulating concentration of 1,25(OH)2D3 is tightly regulated by renal 1α-OHase and kept within a relatively narrow range. Several studies have revealed an inverse correlation between 25(OH)D3 plasma levels and the risk of breast cancer (10-12). The risk of breast cancer is more than 5 times as high for women with a plasma 25(OH)D3 concentration of <50 nmol/l than those with a plasma concentration exceeding 150 nmol/l (12).
The liganded vitamin D receptor (VDR) and the retinoid X receptor are bound to vitamin D-responsive elements by 1,25(OH)2D3 in the CYP24 promoter region. So according to its role as a negative feedback enzyme, 24-OHase expression is induced (13). Active 1,25(OH)2D3 is metabolized by 24-OHase to less active products, which compete with 1α-OHase for their common substrate, 25-(OH)D3. Thus the risk of potentially harmful hypercalcemic side-effects of vitamin D are reduced (14).
Relatively low levels of catabolic 24-OHase activity have been found in extrarenal tissue (11, 15, 16).
Different splice variants of a given protein can display different biological functions and may cause tissue-specific variations in healthy cells. A number of studies have shown that alternative splicing of 24-OHase occurs in different human cells (e.g. breast cancer, endometrial and myelomonocytic cells) (17-19). These splice variants may lead to a catalytically dysfunctional protein.
In this study, the expression of 24-OHase in the human breast cancer cell line MCF-7 and benign breast cell line MCF-10F, as well as in benign and malignant breast tissue, was investigated.
Materials and Methods
Malignant tumor and benign tissue was obtained from resection of Caucasian female patients who had undergone surgery for invasive breast cancer or benign tumors, such as fibroadenoma, at the University of Schleswig-Holstein, Campus Luebeck, with patient consent and local ethical consent. The mean age at primary tumor diagnosis (63 years (range 36-76 years)) and histology were validated from histopathology reports and patient medical records. A sample of breast tissue was excised from the relevant frozen surgical sample which had been stored in liquid nitrogen.
Cell culture. The human benign breast cell line MCF-10F (ATCC No. CRL-10318) and breast cancer cell line MCF-7 (ATCC No. HTB-22) were purchased from the European Collection of Cell Culture (Wiltshire, UK). The MCF-7 cells were maintained in RPMI-1640 medium (Gibco BRL, Karlsruhe, Germany) supplemented with 25 mM Hepes, 1% L-glutamine and 10 % fetal bovine serum (FBS) and the MCF-10F cells in a 1:1 mixture of Dulbecco's modified Eagle's medium and Ham's F12 medium supplemented with 20 ng/ml epidermal growth factor (EGF), 100 ng/ml cholera toxin, 0.01 mg/ml insulin, 500 ng/ml hydrocortisone and 0.04 mM final calcium concentration 5% horse serum (Invitrogen, Karlsruhe, Germany).
RNA isolation. The total RNA from cell culture and homogenized healthy and malignant breast tissue samples was extracted with Trizol (Invitrogen) according to the manufacturer's instructions. The integrity of the isolated RNA was verified by 1% agarose gel electrophoresis and it was quantified spectrophotometrically at OD 260/280 nm.
Reverse transcription (RT) and real time PCR. The synthesis of cDNA from 2 μg of the total RNA was performed using Super Script-II reverse transcriptase (Invitrogen), the RT reaction mixture was diluted 1:5 and 2 μl of the RT reaction mixture was used as the template for real-time PCR with 0.5 mmol/l primers (TBP forward 598-618 5′-CCA CTC ACA GAC TCT CAC AAC-3′, reverse 5′-CTG CGG TAC AAT CCC AGA ACT-3′; 24-OHase forward: 5′-GCA GCC TAG TGC AGA TTT-3′, reverse: 5′-ATT CAC CCA GAA CTG TTG-3′). After adding 12.5 μl of Platinum® SYBR® Green qPCR SuperMix-UDG (Invitrogen), the volume was adjusted to 25 μl with nuclease-free water. The samples were amplified by the DNA Engine Opticon 2™ (Biorad; Hercules, CA, USA) System and PCR was performed by an initial denaturising step at 50°C for 2 min and 95°C for 2 min followed by 50 cycles with a denaturising step at 95°C for 15 s, primer annealing at 57.3°C (24-OHase) and 60°C (TBP) for 15 s, and an extension phase at 72°C for 15 s for 24-OHase and hypoxanthin-phosphoribosyl-transferase (HPRT).
A melting curve was generated after 50 cycles for the final PCR product of all the genes investigated by decreasing the temperature to 65°C for 15 s followed by a gradual increase in temperature to 95°C. During the gradual heating process the fluorescence was measured at increments of 0.2°C. The relative 1α-OHase gene expression between the benign and malignant tissue samples was obtained by normalizing to MCF7 as the calibrator and TBP as the housekeeping gene. The fold change was determined with the formula: efficiency target gene^ (calibrator investigated gene-sample investigated gene)/efficiency housekeeping gene^(calibrator housekeeping gene-sample housekeeping gene). The experiments were performed in triplicate for each gene.
Statistical evaluation. Statistical analysis of the real-time PCR results was performed using Student's t-test.
Western blot. The cells were maintained under standard cultivation conditions, washed twice with PBS and lysed in a sample buffer (125 mM Tris, 30% glycerine, 8% SDS, pH 6.8). The frozen samples of benign and malignant breast tissues were homogenized with Ultra Turrax T25 in tris buffered saline (TBS) containing 0.1% Triton® X-100 and 1mM PMS buffer and lysed in the aforementioned sample buffer. Ten μg of the protein lysates were separated on 12.5% SDS PAGE under reducing conditions and transferred to a nitrocellulose membrane (Optitran BA-S 85; Schleicher Schuell, Dassel, Germany). To eliminate non-specific binding sites, the membranes were blocked overnight at 4°C with blocking agent (PBST with 5% non-fat powdered milk). The membranes were then incubated with primary antibodies against human 24-OHase (Santa Cruz Biotechnologies, Heidelberg, Germany) at a dilution of 1:5,000 for the tissue protein samples and 1:2,500 for the cell culture protein lysates overnight at 4°C.
The secondary antibodies conjugated to horseradish peroxidase (donkey anti-goat IgG; Calbiochem, Darmstadt, Germany) were added at a dilution of 1:10,000. The obtained signals were visualized using the enhanced chemiluminescence (ECL) detection system (Amersham Biosciences, Freiburg, Germany) and as loading control compared to β-actin.
Results
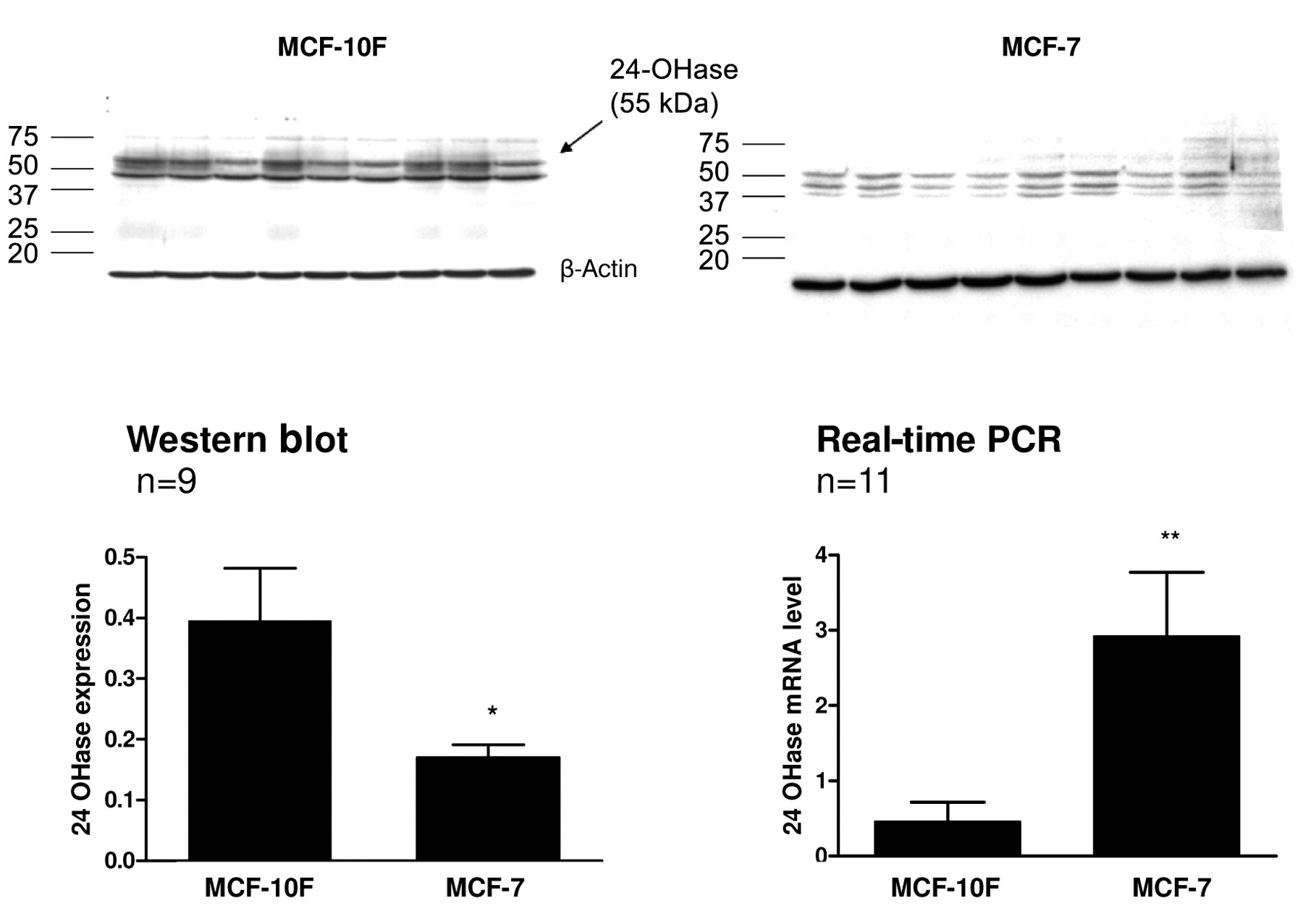
The expression of 24-OHase mRNA was found to be 7.23 times higher in the MCF-7 compared to the MCF-10F cells. In contrast, the expression of 24-OHase protein in the malignant MCF-7 cells, when compared with the benign MCF-10F cells, was lower by 5% to 42.9% (p<0.05) (Figure 1).
In the Western blot, in both cell lines the polyclonal antibody detected a signal at 55 kDa that corresponded to the size of normal 24-OHase protein. A second band was detected at 46 kDa in the MCF-7 as well as in the MCF-10 cells. In the MCF-7 cells, these signals were weaker, which indicated a lower level of active enzyme in these cells. In the MCF-7 cells, there was at least one smaller variant at 42 kDa (Figure 1).
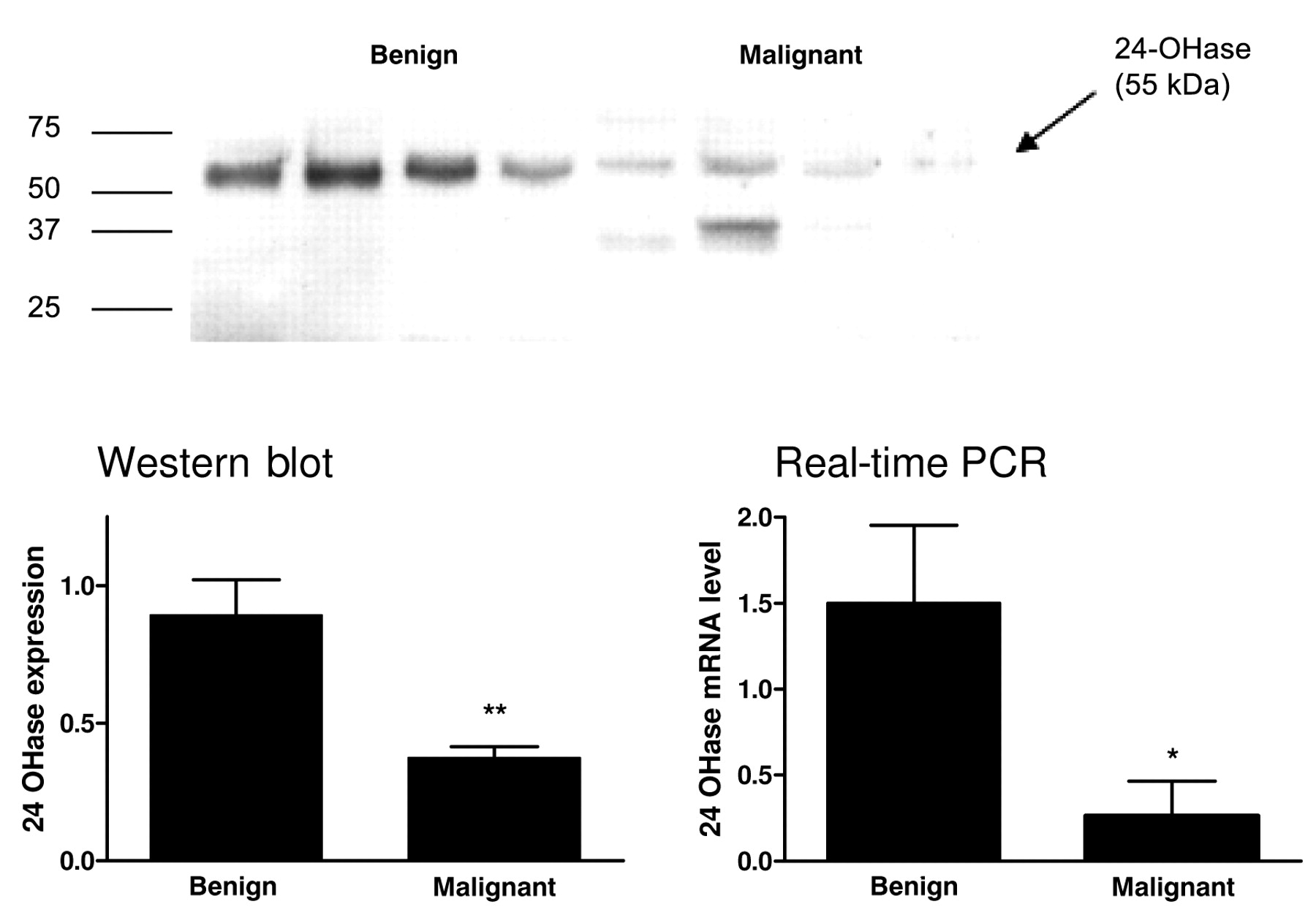
The studies of the malignant and benign breast tissue revealed similar results (Figure 2). The expression of 24-OHase in the malignant tissue decreased, compared to the benign tissue, by 58% to 42% (p<0.001). At the mRNA level the 24-OHase expression in the cancer was reduced compared to the benign tissue by 83% (p<0.001). The protein determination through Western blot continuously showed a distinctive band of 55 kDa in the benign tissue and further bands could not be detected. A band of 55 kDa was only detected in some of the breast cancer tissue samples. However, a weak band of 42 kDa was also detected in some of the samples.

The expression of (A) 24-OHase protein (Western blot) and (B) 24-OHase mRNA (quantitative real-time PCR) in human breast cell lines (n=9). The data are presented as relative mRNA fold change of malignant compared to benign cells, calculated according to the calibrator normalized method and standardized to the endogenous control (*p<0.05, **p<0.001).
Discussion
CYP24 is one of the most sensitively regulated target genes for 1,25(OH)2D3 (20). The correlation in expression between 1α-OHase, VDR and 24-OHase in benign breast tissue provided evidence of localized synthesis of 1,25(OH)2D3. An increased expression of 24-OHase in breast tumors was previously assumed to be a response to the enhanced localized production of 1,25(OH)2D3 (19).
In the present study in the cell lines, a band was found in the Western blot at 56 kDA, which matched the wild-type of the enzyme. In the MCF-7 cells, aside from a weaker main band, we found a number of additional bands. This could indicate that, on the one hand, the wild-type enzyme existed to a lower extent, while on the other hand, there are a number of splice variants which do not show any functional activity. Due to the lack of activity measurements, this can only remain a hypothesis that supports the observation that lower synthesis as well as a lower level of 1,25(OH)2D3 occurs in breast cancer (21, 22).
A higher mRNA level was shown in the RT-PCR of the malignant cells than the benign cells. This could have indicated more functionally inactive variants. Wu et al. detected an intron 2-containing splice variant, which represented a catalytically inactive enzyme. In addition, single nucleotide polymorphism (SNP) in an intron as well as different splice patterns between exons 9 and 11 in the 24-OHase gene were detected (22). An intron-containing variant could reduce the expression of normal 24-OHase enzymes as non coding RNA (ncRNA) with a regulatory effect. The intron 2-containing splice variant described by Ren et al. was detected in macrophages (19) and encoded a non-functional 24-OHase enzyme. The expression of further intron-containing ncRNA does not appear unlikely. Such expression in macrophages of the inflammatory infiltrate of a carcinoma would also explain why the reduced protein expression was limited to malignant mammary tissue.

The expression of (A) 24-OHase protein (Western blot) and (B) 24-OHase mRNA (quantitative real-time PCR) in human breast tissue samples. The data show relative mRNA fold change of malignant compared to healthy breast tissue, calculated according to the calibrator normalized method and standardized to the endogenous control (*p<0.05, **p<0.001).
In previous studies, we cloned a 2.5 kB cDNA splice variant which was 200 bp shorter than the wild-type and corresponded to a 36 kDa-band. In the present study, additional bands were also found in the MCF-7 and -10 cells, measuring between 56 and 40 kDa. These bands were detected very constantly, so that if further splice variants are assumed, accumulation could take place through the inactive variants. Splice variant and wild-type peptides may be in direct competition with each other for substrate binding.
In the tissue as in the cell lines, a clearly more specified band was detected in the benign tissue than in the malignant. In some of the cancer tissue samples, further bands at about 37 kDa were detected. These comply with the splice variants cloned by Ren et al. (19). Furthermore, compared to the malignant tissue, an increase in protein level was found in the benign tissue. The increased mRNA expression in the benign tissue was even clearer although the examined tissue was very heterogeneous. Moreover, the results complied with the results of Anderson et al. regarding the mRNA level, which showed reduced expression in mammary carcinoma compared to benign breast tissue (23).
The measurement of activity related to the observations and any functional consequences should also be specified. Townsend et al. showed activity measurements, contrary to the present data, in that the RNA in malignant tissue, as well as the expression of the protein was highly regulated (24). A stronger degradation of calcitriol could support carcinogenesis. In malignant cells, a significant increase of 24-OHase RNA was detected (24), which was consistent with the present results and in accordance with the research of Tanner et al. (25), who declared 24-OHase to be a potential oncogene after DNA array and positional cloning analyses of genomic amplification.
Examination of the autocrine or paracrine effects of 1,25(OH)2D3 has become more and more interesting in connection with the protective and growth reducing attributes of the hormone. It is uncertain whether breast tissue has lower synthetic 1α-OHase activity compared to other cells, which would explain the higher potential for malignant degeneration. Among women more exposed to UV-radiation, lower activity of 1α-OHase is partially neutralized by a higher supply of vitamin D3 (26). Whether decreased 24-OHase expression could be explained by decreased serum level of vitamin D3, or whether a change in activity in the enzyme is explained by variants, has to be examined in further studies.
Acknowledgements
We would like to thank Birte Münchow for her support.
- Received January 29, 2009.
- Revision received May 21, 2009.
- Accepted May 27, 2009.
- Copyright© 2009 International Institute of Anticancer Research (Dr. John G. Delinassios), All rights reserved
References
In this issue





{kind=link}
{kind=link}
Jump to section
Related Articles
Cited By...
- The Relative Expression of ER{alpha} Isoforms ER{alpha}66 and ER{alpha}36 Controls the Cellular Response to 24R,25-Dihydroxyvitamin D3 in Breast Cancer
- Alternative Splicing in the Cytochrome P450 Superfamily Expands Protein Diversity to Augment Gene Function and Redirect Human Drug Metabolism
- Antineoplastic effects of 1,25(OH)2D3 and its analogs in breast, prostate and colorectal cancer
- Serum 25-Hydroxyvitamin D Levels in Patients with Vulvar Cancer
- Correlation of Prostaglandin Metabolizing Enzymes and Serum PGE2 Levels with Vitamin D Receptor and Serum 25(OH)2D3 Levels in Breast and Ovarian Cancer
- Vitamin D, Calcidiol and Calcitriol Regulate Vitamin D Metabolizing Enzymes in Cervical and Ovarian Cancer Cells