


Abstract
The high incidence of prostate cancer and lack of an effective, long-term treatment for metastatic disease highlights the need for more potent non-calcemic vitamin D analogs as potential alternative or combinational prostate cancer therapies. Among the analogs, 19-nor-1α,25-dihydroxyvitamin D2 (19-nor-1α,25(OH)2D2) known as paricalcitol or Zempler, has less calcemic effects and an equipotential activity as 1α,25-dihydroxyvitamin D3 (1α,25(OH)2D3) in several in vivo and in vitro systems. It was recently demonstrated that a modified analog of paricalcitol, 19-nor-2α-(3-hydroxypropyl)-1α,25-dihydroxyvitamin D3 (MART-10) compared to 1α,25(OH)2D3 was more effective in inhibiting proliferation of an immortalized normal prostate cell line (PZ-HPV-7) (1,000-fold) and invasion of PC-3 prostate cancer cells (10-fold). In this study, the effects of MART-10 and 1α,25(OH)2D3 on proliferation, vitamin D receptor transactivation, vitamin D-binding protein (DBP) binding, CYP24A1 (24-OHase) substrate hydroxylation kinetics, and induction of CYP24A1 gene expression were compared in an androgen-dependent prostate cancer cell model, LNCaP. The results demonstrated that MART-10 was 1,000-fold more active than 1α,25(OH)2D3 in inhibiting LNCaP cell proliferation. MART-10 was more active than 1α,25(OH)2D3 in up-regulating a vitamin D receptor-responsive Luciferase construct and inducing CYP24A1 gene expression in LNCaP prostate cancer cells. In addition, MART-10 has a lower affinity for DBP and less substrate degradation by CYP24A1 compared to 1α,25(OH)2D3, indicating that MART-10 has more bioavailability and a longer half-life. Thus, these data suggest that MART-10 may be a potential candidate as a therapeutic agent for prostate cancer, especially for patients who fail in conventional therapies.
Prostate cancer is a commonly diagnosed malignancy as well as the second leading cause of cancer deaths among men in the United States (1). More than 40,000 men die from prostate cancer each year, accounting for one in six male deaths in the US alone. Worldwide, prostate cancer ranks third in cancer incidence and sixth in cancer mortality among men. More than 670,000 men a year are diagnosed with prostate cancer, accounting for one in nine of all new cancer cases in males. It is now clear that prostate cell growth is regulated not only by androgens but also by another steroid hormone, vitamin D, through vitamin D receptor (VDR)-mediated signaling pathways (2). The incidence of prostate cancer and lack of effective, long-term treatment for metastatic disease highlights the need for alternative therapies, such as vitamin D or vitamin D analog-based therapy, as potential options for prostate cancer treatment.
Several steps in prostate cancer progression can be regulated by the active form of vitamin D, 1α,25-dihydroxyvitamin D3 (1α,25(OH)2D3). 1α,25(OH)2D3 can inhibit epithelial or metastatic cancer cell growth by inducing cell cycle arrest, differentiation or apoptosis (3) or reducing tumor metastasis through regulation of proteases (4). The antiproliferative effect of 1α,25(OH)2D3 in cancer cells is mediated through VDR, a member of the nuclear receptor superfamily that includes receptors for steroid hormones (e.g. androgen and estrogen) as well as for thyroid hormone and retinoids (5). VDR binds 1α,25(OH)2D3 with high affinity and specificity, recruits retinoid X receptor (RXR) to form a heterodimer complex and binds to vitamin D-response elements (VDRE) in the promoter region of vitamin D-regulated genes (6). The C-24 hydroxylation of 1α,25(OH)2D3, catalyzed by 25-hydroxyvitamin D 24-hydroxylase (24-OHase or CYP24A1), is the major pathway by which cells terminate 1α,25(OH)2D3 activity via side-chain cleavage. CYP24A1 contains a VDRE in its proximal promoter and is up-regulated by its own substrate, 1α,25(OH)2D3 (7). 1α,25(OH)2D3-mediated induction of CYP24A1 gene is considered the most sensitive indicator of 1α,25(OH)2D3 biological effects in cells.
Over the last decade or two, investigators have been synthesizing analogs of 1α,25(OH)2D3, seeking to reduce the calcemic properties while retaining its antiproliferative properties (8, 9). Although 1α,25(OH)2D3 exhibits the ability to inhibit cell growth in prostate cancer, clinical use of high concentrations of 1α,25(OH)2D3 is precluded due to the unacceptable side-effects of elevated serum and urine calcium levels (10). Thus, much effort has been expended in designing analogs of 1α,25(OH)2D3 that retain the antiproliferative effects of 1α,25(OH)2D3, but are less calcemic when administered in vivo (8, 9). One such vitamin D analog that has shown promise is the 19-nor-1α,25(OH)2D2 analog (paracalcitol) synthesized by DeLuca and colleagues (11). 19-nor-1α,25(OH)2D2 has properties similar to 1α,25(OH)2D3 in inducing vitamin D target genes and suppressing parathyroid hormone secretion in hemodialysis patients with secondary hyperparathyroidism, without inducing hypercalcemia or hyperphosphatemia (12). The mechanisms responsible for the analogs being less calcemic are not fully understood. However, cellular uptake, metabolism and tissue-specific recruitment of co-activators and/or co-repressors due to conformation changes in VDR/RXR heterodimer have been suggested (8, 9). The effects of A-ring modifications of 19-nor-1α,25(OH)2D3 at the carbon-2 proximal to the 1-alpha hydroxyl group in prostate cells were investigated in order to develop more potent anticancer agents with less calcemic side-effects (13). It was found that one of the 19-nor-1α,25(OH)2D3 analogs, 19-nor-2α-(3-hydroxypropyl)-1α,25(OH)2D3 (MART-10), was approximately 1,000-fold more potent than 1α,25(OH)2D3 in inhibiting cell proliferation of PZ-HPV-7 normal prostate cells. These data suggest that MART-10 has higher biological activity and is more potent than 1α,25(OH)2D3.
In this report, MART-10 was investigated as a candidate therapeutic agent for treating prostate cancer by comparing its effects with those of 1α,25(OH)2D3 on proliferation, VDR transactivation, vitamin D-binding protein (DBP) binding, 24-OHase substrate hydroxylation kinetics, and induction of 24-OHase gene expression in an androgen-dependent prostate cancer cell model, LNCaP.
Materials and Methods
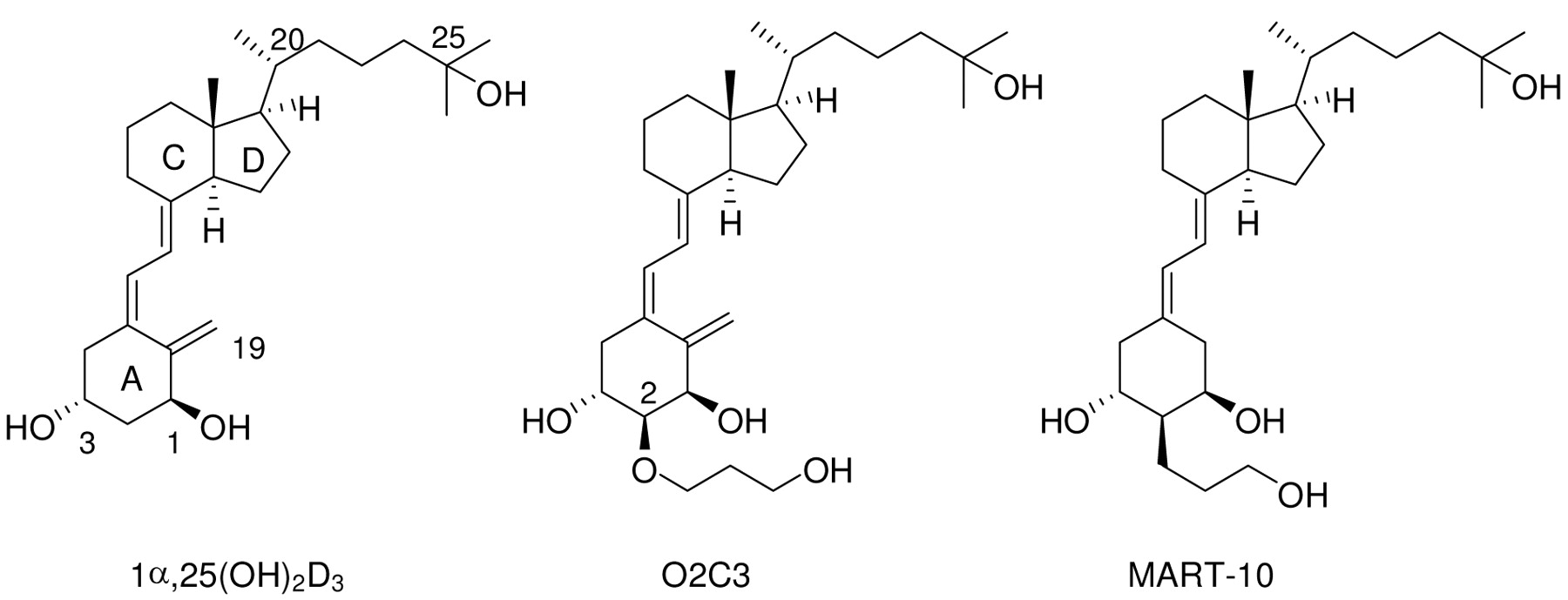
Vitamin D compounds. 1α,25(OH)2D3 was a generous gift from Dr. M. Uskokovic. 19-Nor-2α-(3-hydroxypropyl)-1α,25(OH)2D3 (MART-10) (14) and 2α-(3-hydroxypropoxy)-1α,25(OH)2D3 (O2C3) (17) were synthesized in the laboratory of Dr. A. Kittaka (14).
Cell culture. The LNCaP cell line was obtained from the American Type Culture Collection (Manassas, VA, USA). LNCaP cells were grown in RPMI media (SIGMA, St Louis, MO, USA) supplemented with 5% FBS (GINCOBRL, Gaitherburg, MD, USA). The medium was changed three times per week.
Cellular proliferation assay. The cell counting method using a hemocytometer was performed to assess the antiproliferative activities of MART-10 and 1α,25(OH)2D3. Initially, LNCaP cells were plated at 5,000 cells per cm2 in 35 mm dishes. Two days after the initial plating, the cells were treated with ethanol vehicle, 1α,25(OH)2D3 at 10-9, 10-8, 10-7 and 10-6 M, or MART-10 at 10-12, 10-11, 10-10 and 10-9 M in FBS-supplemented medium. The treatment was repeated two and four days later. After three doses, the medium was removed from the cultures. The attached cells were then trypsinized for 5 minutes with trypsin/EDTA at 37°C and then neutralized with FBS-supplemented growth medium. Each sample of cells was spun down and resuspended in equal volumes of growth medium. Triplicate aliquots were applied to a hemocytometer for cell counting of four different fields under a light microscope and then averaged. The results are expressed as the percentage of control counts.
Real-time qPCR analysis. LNCaP cells were cultured and treated with ethanol vehicle, 1α,25(OH)2D3 or MART-10 at 10-9 or, 10-8, and 10-7 Μ in FBS-supplemented medium for 24h. Total RNA was extracted using the SV Total RNA Isolation kit (Promega, Madison WI, USA). For each sample, cDNA was generated with 2 μg of total RNA using Superscript RNAase H-(Invitrogen, Carlsbad, CA, USA) with random hexamer primers. For each real-time qPCR reaction, 20 ng of single-stranded cDNA was mixed with 2X SYBR Green PCR Master Mix (Applied Biosystems, Foster City, CA, USA) and an optimal concentration of sequence specific primers. The PCR was performed with an ABI Prism 7500 (Applied Biosystems) as follows: initial denaturation at 95°C for 10 min, and 40 cycles of denaturation at 95°C for 30 s, annealing/extension at 60°C for 1 min. Human sequence-specific primers were as follows: CYP24A1, forward 5′-GGC CTG GAT GTC GTA TTT GC-3′ and reverse, 5′-TGA ACG TTG GCT TCA GGA GAA-3′. To normalize the amount of sample cDNA added to each reaction, Taqman PDAR eukaryotic 18S rRNA, (Applied Biosystems) was used as the endogenous control. Relative quantitation of gene expression was calculated using the ΔΔCt method (User Bulletin publication #2, Applied Biosystems).
VDR transactivation. Cells at 60-85% confluence were transiently transfected in triplicate with pVDRE-Luc (0.2 μg per well) by Lipofectamine Plus reagent according to the manufacturer's protocol (Invitrogen, San Diego, CA, USA) (15). After transfection, cells were fed DMEM with charcoal-stripped 10% FCS, and incubated at 37°C overnight. The next day cells were treated with ethanol vehicle, 1α,25(OH)2D3 MART-10 at 10-10 or , 10-9 and 10-8 M. After 24 h, the cells were harvested and cell extracts were prepared for luciferase activity measurement. Luciferase activity is expressed as percentage of ethanol-treated control cells after normalization for protein concentration.
Vitamin D binding assay. The binding affinity of 1α,25(OH)2D3 and MART-10 for DBP was determined by the displacement of [3H]-25(OH)D3 from rat serum DBP by 1α,25(OH)2D3 or MART-10 as described previously (16). Briefly, 10 μL of [3H]-25(OH)D3 (containing 2,500 cpm of radioactivity) were added to duplicated tubes containing increasing concentrations of 1α,25(OH)2D3 (0.0781-5.0 μM) or MART-10 (1.562-50.0 μM) in 50 μL ethanol, followed by adding 500 μL of rat serum pre-diluted with barbital-acetate buffer, pH. 8.6, with BSA as described (16). The mixtures were vortex-mixed and incubated overnight at 4°C. One hundred microliters of dextran-charcoal (0.03% dextran T-70, 0.3% Norit GSX Charcoal) in barbital-acetate buffer, pH.8.6, were added to the mixture, followed by centrifugation at 1000 × g for 15 min at 4°C to separate bound from free [3H]-25(OH)D3. The supernatant was transferred to counting vials and the radioactivity measured with a liquid scintillation counter.
Metabolism of MART-10 by CYP24A1 (24R-OHase) in a reconstituted cell-free system. CYP24A1-dependent metabolism of MART-10 and 2α-(3-hydroxypropoxy)-1α,25(OH)2D3 (O2C3) were measured in a reconstituted system containing the membrane fraction as described previously (17). The activities were measured under the following conditions: 0.05 μM human CYP24A1, 2.0 μM ADX, 0.2 μM ADR, 0 to 5.0 μM MART-10 or O2C3 in 100 mM Tris-HCl (pH 7.4) and 1 mM EDTA at 37°C. The reaction was initiated by the addition of NADPH at a final concentration of 0.5 mM. Aliquots of the reaction mixture were collected at various time-intervals and extracted with 4 volumes of chloroform-methanol (3:1). The organic phase was recovered and dried under reduced pressure. The resultant residue was dissolved in acetonitrile and applied to HPLC under the following conditions: column, YMC-Pack ODS-AM (5 μm) (4.6 mm × 300 mm) (YMC Co., Kyoto, Japan); UV detection, 254 nm for MART-10, and 265 nm for O2C3; flow-rate, 1.0 mL min-1; column temperature, 40°C; mobile phase, a linear gradient of 20-100% acetonitrile aqueous solution for the first 25 min followed by 100% acetonitrile for 10 min for the isolation and identification of MART-10, O2C3 and their metabolites.
Statistics. The data of each group were compared by the Student's t-test using Microsoft Excel 2007. P-value <0.05 was considered as indicating significant difference.
Results
MART-10 has increased inhibitory effects on cellular proliferation of LNCaP prostate cancer cells. It has been previously demonstrated that 19-nor-1α,25(OH)2D2 has similar activity to 1α,25(OH)2D3 in inhibiting the proliferation of human prostate cancer cell lines in vitro, including LNCaP, an androgen-dependent cell line (18). Thus, the inhibitory effects of MART-10 and 1α,25(OH)2D3 on the proliferation of LNCaP cells were compared. As shown in Figure 1, 1α,25(OH)2D3 inhibited of LNCaP cell proliferation to 99±7, 60±8, 41±1 and 28±5% of the control at 10-9, 10-8, 10-7 and 10-6 M respectively, and MART-10 by 97±8, 62±5, 36±3 and 22±3% at 10-12, 10-11, 10-10 and 10-9 M, respectively. These data indicated that MART-10 is about 1,000-fold more active than 1α,25(OH)2D3 in inhibiting LNCaP cell proliferation.

The dose-dependent effects of 1α,25(OH)2D3 and MART-10 on LNCaP cell proliferation. LNCaP cells were treated with ethanol vehicle or the indicated concentrations of 1α,25(OH)2D3 or MART-10 for one week as described in Materials and Methods, and then trypsinized and cell counted with a hemocytometer. The results are expressed as a percentage of the control of the means±SD of 3 determinations from a representative experiment. The experiment was repeated 3 times with similar results. *P<0.05 compared to the lower doses.
MART-10 enhanced CYP24A1 (24-OHase) mRNA expression in LNCaP prostate cancer cells. The 24-OHase gene is induced by VDR-mediated 1α,25(OH)2D3 transactivation through a VDRE located in its proximal promoter (7). It has been previously demonstrated that CYP24A1 gene expression induced by 1α,25(OH)2D3 and vitamin D3 is a simple and effective way to monitor vitamin D-mediated gene transactivation in prostate cells (19, 20). Thus, the expression of CYP24A1 from LNCaP cells was monitored by real-time quantitative PCR in response to 1α,25(OH)2D3 and MART-10 treatment. As shown in Figure 2, MART-10 induced a higher expression level of CYP24A1 at 10-9 and 10-8 M compared to 1α,25(OH)2D3. However, 1α,25(OH)2D3 had a higher induction of CYP24A1 expression at 10-7 M compared to MART-10.
MART-10 induces VDR transactivation at a lower concentration than 1α,25(OH)2D3. To compare the VDR-activating properties of its natural agonist 1α,25(OH)2D3 with that of MART-10, 1α,25(OH)2D3-responsive luciferase (Luc) reporter plasmid pVDRELuc (15) was transfected into LNCaP cells with ethanol control, or increasing concentrations of 1α,25(OH)2D3 or MART-10 for 24 h. Cells were then harvested and the normalized promoter activity was determined as a read-out for VDR-mediated transcriptional activation. As shown in Figure 3, MART-10 stimulated VDR reporter transactivation activity to 30% over control at 10-10 M (p<0.05), and the stimulation increased to 70% and 270% over the control at 10-9 and 10-8 M, respectively. No stimulation by 1α,25(OH)2D3 was observed at 10-10 and 10-9 M. At 10-8 M 1α,25(OH)2D3, a 210% stimulation was observed. Thus, the data indicate that MART-10 is able to stimulate VDR transactivation at much lower concentrations than 1α,25(OH)2D3, and is at least 10-fold more active than 1α,25(OH)2D3 in stimulating VDR transactivation in LNCaP cells.

The effects of 1α,25(OH)2D3 and MART-10 treatment on endogenous 24-OHase mRNA expression in LNCaP cells. LNCaP cells were treated with ethanol vehicle, or the indicated concentrations of 1α,25(OH)2D3, or MART-10 for 24 h, and then total RNA was prepared and analyzed by relative real-time quantitive PCR. Data are expressed as the mean of triplicate determinations. The data were calculated according to the ΔΔCt method and standardized to an endogenous control, S18. Values represent the percentage change in gene expression relative to ethanol vehicle-treated control LNCaP cells.
MART-10 has a lower binding affinity for DBP than 1α,25(OH)2D3. In order to investigate the bioavailability of MART-10, its ability to bind serum DBP was examined. Figure 4 demonstrates the binding affinities of 1α,25(OH)2D3 and MART-10 for DBP. The dissociation constant (Kd) was defined as the concentration at which 1α,25(OH)2D3 and MART-10 still occupied 50% of the DBP binding sites. The Kd for MART-10 and 1α,25(OH)2D3 were determined to be 17.5 μM and 0.67 μM, respectively. Thus, the data indicate that the binding affinity of MART-10 for DBP is about 25-fold less than 1α,25(OH)2D3. The lower binding affinity for MART-10 will result in a higher concentration of free bioavailable MART-10 in the circulation with its subsequent translocation to various target tissues, including the prostate.

The effects of MART-10 and 1α,25(OH)2D3 on VDR transactivation in LNCaP cells. LNCaP cells tranfected with pVDRELuc reporter construct were treated with ethanol control or the indicated concentrations of 1α,25(OH)2D3 or MART-10 for 24 h, then cells were harvested and assayed luciferase activity. Luciferase activity is expressed as percentage of ethanol-treated control cells after normalization for protein concentration. The data are means±SD of 3 determinations from a representative experiment. Similar results were obtained from two other experiments. *P<0.05 compared to control.

Binding of MART-10 and 1α,25(OH)2D3 to vitamin D binding protein (DBP). The binding affinity of 1α,25(OH)2D3 and MART-10 to DBP was determined by the displacement of [3H]-25(OH)D from rat serum DBP by indicated concentrations of MART-10 and 1α,25(OH)2D3. The results are expressed as percentage of displaced [3H]-25(OH)D (B) over total specific bound of [3H]-25(OH)D (Bo).

Structures of 1α,25(OH)2D3, 2α-(3-hydroxypropoxy)-1α,25(OH)2D3 (O2C3) and 19-nor-2α-(3-hydroxypropyl)-1α,25(OH)2D3 (MART-10).
MART-10 is a very poor substrate for CYP24A1. As described previously, O2C3 has a much higher Km value than 1α,25(OH)2D3, and the kcat/Km value of O2C3 was only 2% that of the 1α,25(OH)2D3 value (17), suggesting that the 3-hydroxypropoxy group at carbon-2α of O2C3 significantly inhibits 24-hydroxylation of O2C3 by CYP24A1. Because MART-10 has the same 3-hydroxypropoxy group at the C-2α position as O2C3, MART-10 may be also more resistant to CYP24A1 degradation than 1α,25(OH)2D3. Therefore, the metabolism of MART-10 by CYP24A1 was compared to O2C3 to determine their potential half-life in circulation. It is noted that MART-10 has a higher Km and a lower kcat value than O2C3, which lead to a significantly lower kcat/Km value than O2C3 (Table I). The data, therefore, suggest that both the addition of 3-hydroxypropyl group at C-2 and the deletion of the methylene group at C-19 in the MART-10 molecule may hinder the binding of MART-10 to the catalytic pocket of CYP24A1, and may also change the conformation of the enzyme molecule thereby affecting its interaction with ADX, an electron donor of CYP24A1, resulting in a very poor 24-hydroxylation and, in turn, much less degradation of MART-10 by CYP24A1.
Discussion
In prostate cancer, androgen deprivation therapy is the mainstay therapeutic option for patients that fail to respond to prostatectomy or radiation. Although three in four men respond to androgen deprivation, the median duration of the response is only about two years. No effective therapies are available at the present time for prostate cancer that becomes resistant to androgen deprivation. Besides androgens, prostate cells also respond to the active form of vitamin D, 1α,25(OH)2D3. Numerous studies have shown that 1α,25(OH)2D3 is a potent inducer of prostate cancer cell differentiation and an inhibitor of prostate cancer cell proliferation, invasiveness and metastasis (2-4, 21). These data strongly indicate that vitamin D-based therapies could be a potential option for prostate cancer treatment when androgen deprivation has failed. However, the use of 1α,25(OH)2D-based therapies is limited by the risk of hypercalcemia and hypercalciuria (10). Therefore, non-calcemic analogs of 1α,25(OH)2D3 with potent antiproliferative activity would be inspiring therapeutic agents (8, 9).
Kinetic parameters of Human CYP24A1 for 2α-(3-hydroxypropoxy)-1α,25(OH)2D3 (O2C3) and MART-10.
MART-10, 19-nor-2α-(3-hydroxypropyl)-1α,25(OH)2D3, was shown to have an antiproliferative potency that is approximately 1,000-fold greater than 1α,25(OH)2D3 in PZ-HPV-7 cells (13). MART-10 was also shown to be about 10-fold more active than 1α,25(OH)2D3 in cell invasion studies using PC-3 prostate cancer cells (13). In the present study, it was demonstrated that MART-10 was more biologically active than 1α,25(OH)2D3 in androgen-dependent LNCaP cells, including cell growth inhibition, VDR transactivation induction and induction of CYP24A1 mRNA expression in LNCaP cells. The enhanced biological activity of MART-10 may be the result of stronger binding affinity with VDR through additional hydrogen bonding. It has been previously shown that vitamin D analogs bound to VDR induce a more constrained structural conformation compared to bound 1α,25(OH)2D3 (22, 23). This constrained conformation of VDR can either hinder or enhance the VDR-RXR heterodimer, in turn affecting the interaction of VDR with other transcription co-regulators. Structural changes to VDR caused by MART-10 could alter the selectivity and magnitude of gene activation (15). More data are needed on the structural binding of MART-10 with VDR to determine its specific transcriptional complex on DNA.
In order to explore other possible explanations for the greater biological activity of MART-10, its bioavailability and deactivation by CYP24A1 were analyzed. It was shown that MART-10, compared to 1α,25(OH)2D3, has a lower binding affinity to DBP, supporting the idea that more free and unbound MART-10 would be available for interaction with VDR to induce its biological activity. It was also found that MART-10 is a poor substrate for CYP24A1 hydroxylation. Hence, MART-10 is deactivated by CYP24A1 at a much slower rate than 1α,25(OH)2D3, resulting in a pooling effect of MART-10. These results could explain the greater activity of MART-10 at lower concentration compared to 1α,25(OH)2D3, however, more research is needed to obtain conclusive data.
The vitamin D-based analog paricalcitol, or 19-nor-1α,25(OH)2D2, has been demonstrated to be safe and effective within a wide dosing range (12) and has been approved for clinical human use. These results demonstrated that a modified 19-nor-1α,25(OH)2D2 analog, MART-10, has higher VDR transactivation activity, antiproliferative ability and CYP24A1 induction activity than 1α,25(OH)2D3 in androgen-dependent LNCaP prostate cancer cells. Moreover, MART-10 binds more weakly to DBP and is more slowly metabolized by CYP24A1 compared to 1α,25(OH)2D3. Thus, MART-10 may be a potential candidate as a therapeutic agent for treating prostate cancer, especially for prostate cancer that has failed in conventional therapies.
Acknowledgements
The authors would like to thank Dr. Carsten Carlberg for providing pVDRE-Luc plasmid. This study was supported in part by Grants-in-Aid for Scientific Research (C) from the Japan Society for the Promotion of Science (No. 17590012 and 19590016 to AK).
Footnotes
-
↵# Current address: Department of Medicine, Section of Andrology, Center for Molecular Medicine, Karolinska Institute, Stockholm, Sweden
- Received January 29, 2009.
- Revision received March 30, 2009.
- Accepted May 5, 2009.
- Copyright© 2009 International Institute of Anticancer Research (Dr. John G. Delinassios), All rights reserved
References
In this issue





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Jump to section
Related Articles
Cited By...
- MART-10, a 1{alpha},25(OH)2D3 Analog, Potently Represses Metastasis of ER+ Breast Cancer Cells with VEGF-A Overexpression
- Paricalcitol Enhances the Chemopreventive Efficacy of 5-Fluorouracil on an Intermediate-Term Model of Azoxymethane-Induced Colorectal Tumors in Rats
- The Roles of Cytochrome P450 Enzymes in Prostate Cancer Development and Treatment