


Abstract
Background: In this study, the effect of novel taxane SB-T-1216 and paclitaxel on sensitive MDA-MB-435 and resistant NCI/ADR-RES human breast cancer cells was compared. Materials and Methods: Cell growth and survival were evaluated after 96-hour incubation with tested concentrations of taxanes. The effect on the formation of microtubule bundles was assessed employing fluorescence microscopy and on the cell cycle employing flow cytometric analysis. The activity of caspases was assessed employing commercial colorimetric kits. Results: The IC50 (concentration resulting in 50% of living cells in comparison with the control) of SB-T-1216 in sensitive cells was 0.6 nM versus 1 nM for paclitaxel. However, the IC50 of SB-T-1216 in resistant cells was 1.8 nM versus 300 nM for paclitaxel. Both SB-T-1216 and paclitaxel at death-inducing concentrations induced the formation of microtubule bundles in sensitive as well as resistant cells. Cell death induced in sensitive and resistant cells by paclitaxel was associated with the accumulation of cells in the G2/M phase. On the contrary, cell death induced by SB-T-1216 took place without the accumulation of cells in the G2/M phase but with a decreased number of G1 cells and the accumulation of hypodiploid cells. Both SB-T-1216 and paclitaxel activated caspase-3, caspase-9, caspase-2 and caspase-8 in sensitive as well as resistant cells. Conclusion: Cell death induced by both paclitaxel and novel taxane SB-T-1216 in breast cancer cells is associated with caspase activation and with the formation of interphase microtubule bundles. Novel taxane SB-T-1216, but not paclitaxel, seems to be capable of inducing cell death without the accumulation of cells in the G2/M phase.
Classical taxanes, such as paclitaxel (Taxol®) and docetaxel (Taxotere®), have been successfully used in the therapy not only of breast and ovarian cancer but also of other types of cancer. Paclitaxel was originally isolated from the bark of the Pacific Yew (Taxus brevifolia) while docetaxel is a semisynthetic taxane (1-5).
Microtubules are polymers of tubulin heterodimers containing α and β subunits (6). Microtubules comprise one of the major components of the cytoskeleton of cells and are involved in many cellular processes including mitotic division (7). At the onset of mitosis, cytoplasmic microtubules undergo dramatic rearrangement to form mitotic spindles. Regular bipolar spindles play a pivotal role in the precise segregation of chromosomes. On the other hand, aberrant multipolar spindles are not compatible with cell viability. Most cells derived from defective mitosis are supposed to undergo apoptosis (8). Taxanes are known as mitotic poisons due to their binding to the β subunit of the tubulin heterodimer. This binding accelerates the polymerization of tubulin, stabilizes microtubules and inhibits their depolymerization (6, 9-11). Thus the interaction of taxanes with microtubules results in the formation of microtubule bundles in interphase cells and the formation of asters instead of mitotic spindles during mitosis. In this way, taxanes are thought to block progression through the M-phase of the cell cycle (12, 13). However, the relationship of the mitotic arrest to the induction of cell death by taxanes is unclear (14-16).
Novel taxanes, i.e. synthesized taxane analogs, represent a new generation of taxoids. Some of them have been found to be more effective or more suitable than paclitaxel and docetaxel in the treatment of various types of cancer cells. They are particularly more effective in resistant cancer cells (4, 17-21). Thus novel taxanes provide us with a better chance in the therapy of such types of cancer as breast and ovarian. Some novel taxanes have been found to exhibit a significant activity towards microtubules (13, 20).
We have shown previously that the novel taxane SB-T-1216 seems more effective, particularly in resistant breast cancer cells, than paclitaxel (18, 19). In the present study, the effect of the novel taxane SB-T-1216 and paclitaxel on the formation of microtubule bundles and cell cycle progression, as well as on the activity of caspase-3, caspase-9, caspase-2 and caspase-8, were compared in sensitive MDA-MB-435 and resistant NCI/ADR-RES human breast cancer cells.
Materials and Methods
Materials. Paclitaxel was obtained from Bristol-Myers Squibb (Princeton, NJ, USA). SB-T-1216 (18) was synthesized in the laboratory of Professor I. Ojima (Stony Brook, NY, USA). Indocarbocyanate (Cy3)-conjugated monoclonal anti-tubulin antibody (recognizing an epitope in the C-terminal of β-tubulin) from Sigma (St. Louis, MO, USA) was used.
Cells and culture conditions. The human breast carcinoma cell lines MDA-MB-435 and NCI/ADR-RES were obtained from the National Cancer Institute (Frederick, MD, USA). Cells were maintained in a culture medium at 37°C in a humidified atmosphere of 5% CO2 in air. The culture medium was RPMI-1640 medium containing extra L-glutamine (300 μg/ml), sodium pyruvate (110 μg/ml), HEPES (15 mM), penicillin (100 U/ml), and streptomycin (100 μg/ml) as described elsewhere (22) supplemented with 10% fetal bovine serum (M. Kysilka, Brno, Czech Republic). For the experiments, cells were harvested employing trypsin (0.2%) + EDTA (0.02%) in PBS.
Cell growth and survival experiments. Cells maintained in the culture medium were harvested by low-speed centrifugation, washed with the culture medium and then seeded at 1×104 cells/100 μl of medium into wells of a 96-well plastic plate. Cell growth and survival were evaluated after 96 h of incubation in the culture medium without taxane (control) and with a range of concentrations (0.01-100 nM) of SB-T-1216. The number of living cells was determined by hemacytometer counting after staining with trypan blue (23).
Propidium iodide staining analysis. Cells grown in the culture medium were harvested by low-speed centrifugation, washed with the culture medium and seeded at 3×106 cells/15 ml of culture medium into plastic culture dishes for 24-h preincubation allowing cells to attach. The medium was changed to culture medium without taxane (control) and with selected concentrations of paclitaxel (30 nM for MDA-MB-435 cells, 3,000 nM for NCI/ADR-RES cells) or SB-T-1216 (10 nM for MDA-MB-435 cells, 100 nM for NCI/ADR-RES). After the required incubation period (24, 48, 72 h), the cells were harvested by low-speed centrifugation, stained and analyzed as described elsewhere (23).
Immunofluorescence microscopy. For immunofluorescence microscopy, cells were seeded in culture medium onto glass slides for 24-h preincubation allowing cells to attach. The medium was replaced after this time with culture medium without taxane (control) or with selected concentrations of paclitaxel or SB-T-1216. After incubation (24 h) attached cells were rinsed twice with microtubule-stabilizing buffer (MSB: 20 mM 2-(N-morpholino)ethanesulfonic acid adjusted to pH 6.9 with KOH, 2 mM EGTA, 2 mM MgCl2, and supplemented with 4% PEG 6000). Cells were fixed for 20 min at 37°C with 3% formaldehyde in MSB and then extracted for 4 min with 0.5% Triton®X-100 in MSB. After rinsing the glass slides with MSB, cells were incubated with the Cy3-conjugated anti-tubulin antibody (1:500) for 45 min at room temperature. The preparations were mounted in Mowiol 4-88 supplemented with 1 μg/ml 4,6-diamidino-2-phenylindole (DAPI) used to label cell nuclei. Visualization was carried out with a fluorescence A70 Provis microscope (Olympus, Hamburg, Germany) and images were captured and processed using a cooled CCD camera SensiCam (Olympus).
Measurement of caspase-3, caspase-9, caspase-2 and caspase-8 activities. Harvested cells were seeded at 5×106 cells/25 ml of culture medium into plastic culture flasks for 24-h preincubation allowing cells to attach, after which the medium was changed to a relevant test medium. After 24 h of incubation in culture medium without taxane (control) or with paclitaxel or SB-T-1216, the cells were harvested by low-speed centrifugation and analyzed as described elsewhere (16, 24). Commercial colorimetric kits Caspase-3/CPP32 Colorimetric Protease Assay, Caspase-9/Mch6/Apaf-3 Colorimetric Protease Assay, Caspase-8/FLICE Colorimetric Protease Assay from Biosource (Camarillo, CA, USA) and solutions from a Caspase-3 Colorimetric Protease Assay kit (Biosource) combined with the chromogenic caspase-2 substrate from Alexis Biochemicals (Lausen, Switzerland) were used. The total protein content in samples was determined by the bicinchoninic acid assay (25).
Statistical analysis. The statistical significance of differences was determined using Student's t-test. P<0.05 was considered statistically significant.
Results
Effect of taxanes on growth and survival. The effect of paclitaxel on sensitive MDA-MB-435 cells and resistant NCI/ADR-RES cells was tested previously (16). Paclitaxel at a concentration of 3 nM and higher induced death of MDA-MB-435 cells within 96 h of incubation, i.e. the number of living cells after the incubation period was lower than the number of cells of the inoculus. The IC50 (concentration of taxane resulting in 50% of living cells in comparison with the control after 96 h of incubation) of paclitaxel was 1 nM. On the contrary, paclitaxel induced death of NCI/ADR-RES cells at a concentration of 1,000 nM and higher; the IC50 of paclitaxel was 300 nM. Approximately 300-fold higher concentrations of paclitaxel were required to induce death of resistant NCI/ADR-RES cells than of sensitive MDA-MB-435 cells. On the basis of these data, 30 nM paclitaxel for sensitive MDA-MB-435 cells and 3,000 nM paclitaxel for resistant NCI/ADR-RES cells were employed as death-inducing concentrations, i.e. the lowest concentrations with full death-inducing effect (16).

Effect of SB-T-1216 (0.01-100 nM) on the growth and survival of MDA-MB-435 and NCI/ADR-RES cells. Control cells (C) were incubated without SB-T-1216. The cells were seeded at 10×103 cells/100 μl of medium in the well. The number of cells of the inoculum is shown as a dotted line. The number of living cells was determined after 96 h of incubation (see Materials and Methods). Each point represents the mean of 8 separate cultures±SEM.
The effect of SB-T-1216 on the growth and survival of sensitive MDA-MB-435 and resistant NCI/ADR-RES cells was tested at a wide range of concentrations (0.01-100 nM). SB-T-1216 induced death of MDA-MB-435 cells within 96 h of incubation at a concentration of 3 nM and higher. The IC50 of SB-T-1216 for MDA-MB-435 cells was 0.6 nM. In the case of NCI/ADR-RES cells, SB-T-1216 induced death at a concentration of 10 nM and higher. The IC50 of SB-T-1216 for NCI/ADR-RES cells was 1.8 nM (Figure 1). The data showed that only about 3-fold higher concentrations of SB-T-1216 were required to induce death in resistant NCI/ADR-RES cells than in sensitive MDA-MB-435 cells. This shows that SB-T-1216 was much more effective in resistant NCI/ADR-RES cells than paclitaxel was. On the basis of these data, death-inducing concentrations of SB-T-1216, i.e. 10 nM for sensitive MDA-MB-435 cells and 100 nM for resistant NCI/ADR-RES cells were used. The death-inducing concentrations of paclitaxel and SB-T-1216 were used in further studies with MDA-MB-435 and NCI/ADR-RES cells.
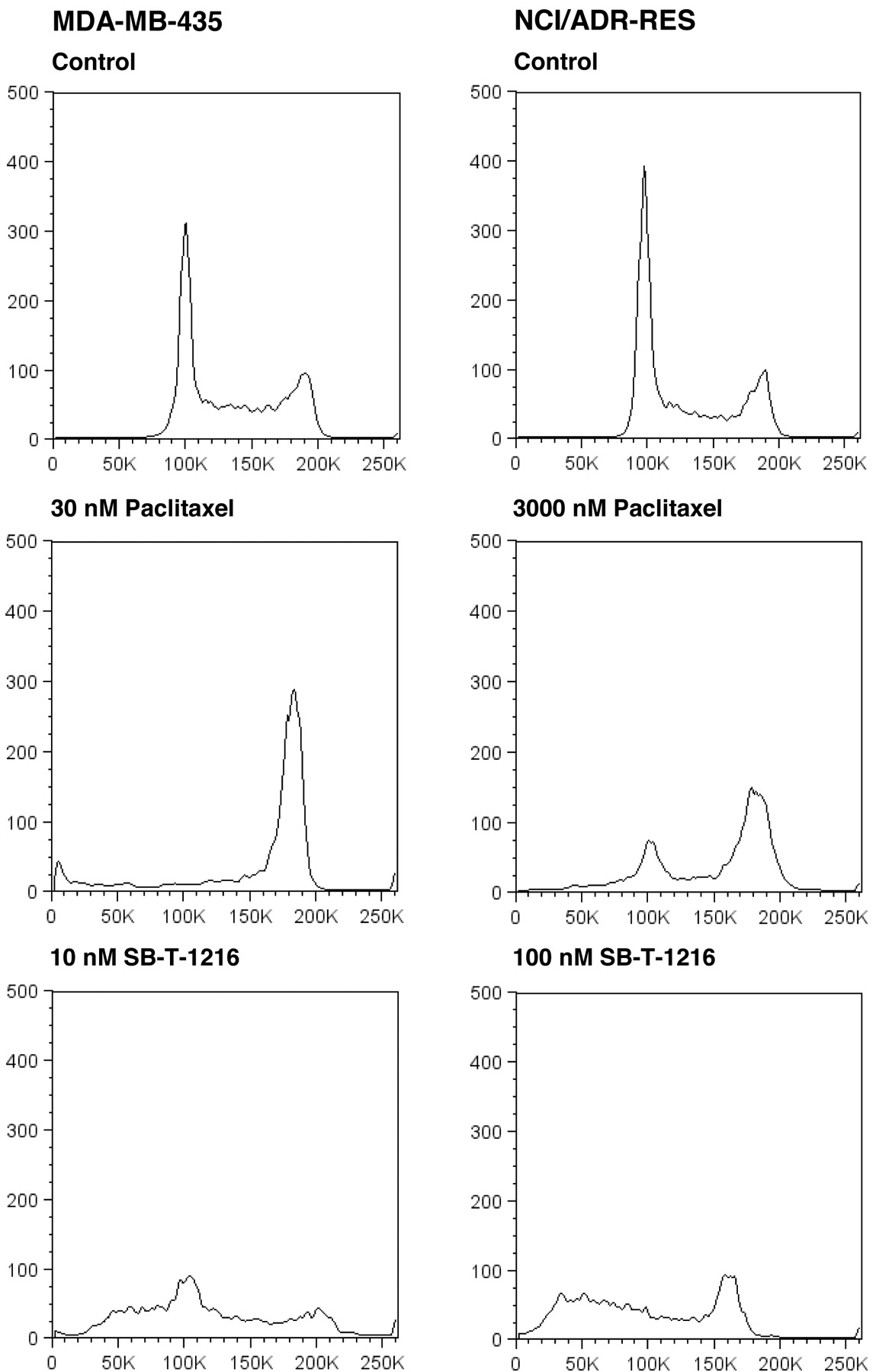
Effect of taxanes on the formation of microtubule bundles. In sensitive MDA-MB-435 cells, paclitaxel at 30 nM as well as SB-T-1216 at 10 nM induced the formation of microtubule bundles within 24 h of incubation (Figure 2). Similarly in resistant NCI/ADR-RES cells, both paclitaxel and SB-T-1216 at 3,000 nM and 100 nM also induced the formation of microtubule bundles within 24 h of incubation (Figure 2).
Effect of taxanes on the cell cycle. Flow cytometric analysis, after propidium iodide staining, showed that the application of paclitaxel at 30 nM resulted in nearly total accumulation of sensitive MDA-MB-435 cells in the G2/M phase of the cell cycle after 24 h of incubation. On the contrary, the application of SB-T-1216 at 10 nM was without any accumulation of the cells in the G2/M phase. The G1 peak significantly decreased and the accumulation of near-G1 hypodiploid cells/particles was apparent (Figure 3).
Similarly, the application of paclitaxel at 3,000 nM resulted in the accumulation of resistant NCI/ADR-RES cells in the G2/M phase after 24 h of incubation. The application of SB-T-1216 at 100 nM was again without accumulation of the cells in the G2/M phase. The G1 peak disappeared and the accumulation of near-G1 hypodiploid cells/particles was apparent (Figure 3). This characteristic effect of 100 nM SB-T-1216 without the accumulation in the G2/M phase was found after 24, 48 and 72 hours of incubation. The G2 peak even disappeared within 72-hour incubation (Figure 4).
Effect of taxanes on caspase-3, caspase-9, caspase-2 and caspase-8 activities. The employed colorimetric assays showed that after 24 h of incubation with paclitaxel or SB-T-1216 at the death-inducing concentrations (30 nM or 10 nM, respectively), the activity of caspase-3 increased significantly in sensitive MDA-MB-435 cells. Caspase-3 activity increased approximately 7.5-fold after the application of paclitaxel (p<0.01) and approximately 5.5-fold after the application of SB-T-1216 (p<0.05). Similarly, caspase-3 activity increased significantly (p<0.01) in resistant NCI/ADR-RES cells after incubation with paclitaxel or SB-T-1216 at the death-inducing concentrations (3,000 nM or 100 nM). The activity of caspase-3 increased approximately 4.5-fold in the case of paclitaxel and approximately 2-fold in the case of SB-T-1216 (Figure 5A).
The activity of caspase-9 also increased significantly in both sensitive MDA-MB-435 (p<0.05) and resistant NCI/ADR-RES (p<0.01) cells after incubation with the death-inducing concentration of paclitaxel or SB-T-1216. The increase was approximately 2.5- to 4-fold for paclitaxel and approximately 3- to 5-fold for SB-T-1216 (Figure 5B).

Effect of paclitaxel and SB-T-1216 at death-inducing concentrations on the formation of interphase microtubule bundles in sensitive MDA-MB-435 and resistant NCI/ADR-RES cells. Control cells were incubated without taxane. After 24 h of incubation, microtubules were detected by fluorescence microscopy following staining with Cy3-conjugated anti-tubulin antibody (red). Cell nuclei were stained with DAPI (blue) (see Materials and Methods). The scale bar represents 10 μm. The data shown were obtained in one representative experiment of two independent experiments.

Effect of paclitaxel and SB-T-1216 at death-inducing concentrations on the DNA histogram of sensitive MDA-MB-435 and resistant NCI/ADR-RES cells. Control cells were incubated without taxane. After 24 h of incubation, the cells were stained with propidium iodide (see Materials and Methods) and analyzed by flow cytometry.
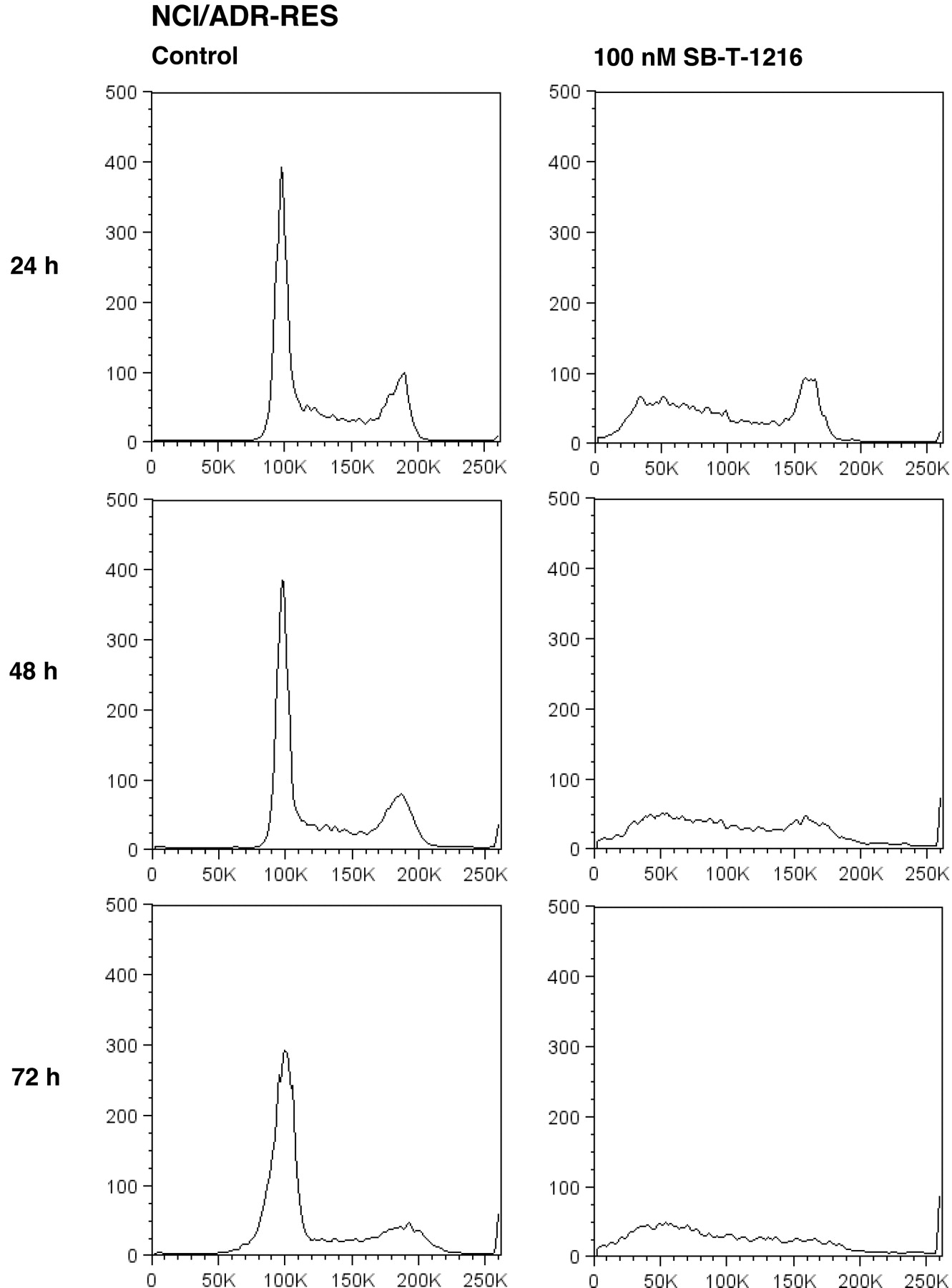
Effect of SB-T-1216 at the death-inducing concentration on the DNA histogram of resistant NCI/ADR-RES cells. Control cells were incubated without taxane. After the incubation period (24, 48, 72 h), the cells were stained with propidium iodide (see Materials and Methods) and analyzed by flow cytometry.

Effect of paclitaxel and SB-T-1216 at death-inducing concentrations on caspase activities in sensitive MDA-MB-435 and resistant NCI/ADR-RES cells. Control cells were incubated without taxane. After 24 h of incubation, the activity of caspase-3 (A), -9 (B), -2 (C) and -8 (D) were measured as the absorbance of the cleaved product of a respective chromogenic substrate at 405 nm, employing a commercial colorimetric kit (see Materials and Methods). Each column represents the mean of 2 experimental values±SEM. *p<0.05, **p<0.01 when comparing with control values.
Concerning caspase-2 activity in sensitive MDA-MB-435 cells, a 7.5-fold increase was detected after incubation with the death-inducing concentration of paclitaxel (p<0.05) and a 4.5-fold increase after incubation with SB-T-1216 (p<0.05). In resistant NCI/ADR-RES cells, we detected a smaller but still statistically significant 2-fold increase of caspase-2 activity after incubation with paclitaxel (p<0.01) and approximately 4.5-fold increase after incubation with SB-T-1216 (p<0.01) (Figure 5C).
A significant (p<0.01) approximately 4.5-fold increase of caspase-8 activity was detected in sensitive MDA-MB-435 cells after incubation with death-inducing concentrations of both paclitaxel and SB-T-1216. In resistant NCI/ADR-RES cells, again a smaller but still statistically significant 1.5-fold increase of caspase-8 activity was detected for paclitaxel (p<0.05) and an approximately 3-fold increase for SB-T-1216 (p<0.01) (Figure 5D).
Discussion
Taxanes are known to modify microtubule dynamics leading to the formation of interphase microtubule bundles and aberrant mitotic spindles. These events are believed to result in blocking of progression through the M-phase of the cell cycle (6, 9, 10, 13). It is also supposed that mitotic arrest could represent a mechanism by which taxanes induce cell death. However, this question has not been clearly answered yet (14-16).
We reported previously that novel taxane SB-T-1216 was more effective than classical taxanes in breast cancer cells (18, 19). Our previous data (16) together with data of the present study (see Figure 1) demonstrate that SB-T-1216 is much more effective than paclitaxel in resistant breast cancer cells and that SB-T-1216 could represent a potentially powerful tool for the treatment of resistant breast cancer. Therefore, we were curious as to the mechanism of the higher efficiency of SB-T-1216 and thus we compared the effect of SB-T-1216 and paclitaxel on the formation of interphase microtubule bundles, cell cycle progression and also on the activity of caspase-3, caspase-9, caspase-2 and caspase-8 in both sensitive MDA-MB-435 and resistant NCI/ADR-RES human breast cancer cells.
Both paclitaxel and SB-T-1216 at death-inducing concentrations induced the formation of interphase microtubule bundles in sensitive MDA-MB-435 cells during 24-h incubation. Similarly paclitaxel and SB-T-1216 induced the formation of interphase microtubule bundles in resistant NCI/ADR-RES cells (see Figure 2). It was shown previously that some novel taxanes can exhibit a significant activity towards microtubuli (11, 13, 20, 21) and that they were able to interact with microtubuli differently than paclitaxel (13). However, it does not seem that SB-T-1216 at death-inducing concentrations exerts differing effect on the formation of microtubule bundles than paclitaxel in tested sensitive and resistant breast cancer cells. Thus our findings suggest that the induction of cell death by both taxanes in breast cancer cells is associated with analogous formation of interphase microtubule bundles.
Cell death induced in both MDA-MB-435 cells and NCI/ADR-RES cells by the death-inducing concentration of SB-T-1216 resulted in a markedly different cell cycle distribution from that of paclitaxel. The effect of paclitaxel was characterized by the accumulation of cells in the G2/M phase. On the contrary, the effect of SB-T-1216 was not associated with the accumulation in the G2/M phase and was characterized by the accumulation of hypodiploid cells (see Figures 3 and 4). These findings support a suggestion that the novel taxane SB-T-1216 at used death-inducing concentrations, but not paclitaxel, induces cell death via a pathway differing from the pathway involving M-phase block. Some previous studies have suggested that taxanes can induce apoptosis independent of mitotic arrest (14, 26-28). This is usually related to lower taxane concentrations while higher taxane concentrations cause cell death connected with mitotic arrest (27-29). Thus, it seems that mitotic arrest does not represent an indispensable prerequisite for taxane-induced cell death. Such suggestion is also supported by the finding that baccatin III, which contains the core taxane ring, can induce apoptosis independent of G2/M arrest (14). We suggest that the potency of SB-T-1216, but not of paclitaxel, to switch on the pathway independent of mitotic arrest in resistant cells can explain, at least partly, the significantly higher efficiency of SB-T-1216 in resistant cells.
The key executioner caspase-3 and upstream caspase-9 were activated in both sensitive MDA-MB-435 and resistant NCI/ADR-RES cells when death was induced by paclitaxel (16) as well as by SB-T-1216 (see Figure 5A and 5B). This fact indicates that the death induced by both taxanes could be related to the mitochondrial pathway of caspase activation as was also suggested by others (30-34). However, significant activation of caspase-2 and caspase-8 was also found in both cell lines after the application of both taxanes at the death-inducing concentration (see Figure 5C and 5D). The role of caspase-2 in apoptosis induction is not clear yet but several lines of evidence point to caspase-2 as being a major player in apoptosis induction (35, 36). Several studies demonstrated caspase-2 activation in various types of cancer cells after apoptosis induction by taxanes (37-39). The involvement of caspase-2 activation in apoptosis of breast cancer cells induced by various stimuli was also demontrated (40-42). Concerning caspase-8 activation, the activation was shown during taxane-induced apoptosis in lymphoma and melanoma cells (38, 39) and during apoptosis induction by HOXA5 in breast cancer cells (40). The activation of caspase-8 suggests that the death receptor pathway of caspase activation could also be somehow involved in cell death induced by taxanes. Considering the activation of all discussed caspases, the functional sequence of the activation of individual caspases remains to be elucidated in order to understand better mechanisms involved in apoptosis induction by taxanes in breast cancer cells.
Taken together, we conclude that cell death induced by both of the tested taxanes, namely paclitaxel and novel taxane SB-T-1216, in breast cancer cells is associated with the activation of several caspases and with the formation of interphase microtubule bundles. However, we suggest that novel taxane SB-T-1216, but not paclitaxel, can induce cell death which is not always directly related to the accumulation of cells in the G2/M phase and thus via a pathway independent of M-phase block. The potency of SB-T-1216 to switch on such a pathway in resistant cells could help to explain its significantly higher efficiency in them.
Acknowledgements
This work was supported by grant NR9426-3/2007 from the IGA, Ministry of Health of the Czech Republic, by grant 301/09/0362 from the Grant Agency of the Czech Republic, by grant 204/05/H023 from the Grant Agency of the Czech Republic, a grant from the National Cancer Institute, USA (CA103314 to I.O.) and a Faculty Development Award from the New York State Office of Science, Technology & Academic Research (to I.O.).
- Received December 23, 2009.
- Revision received May 27, 2009.
- Accepted June 12, 2009.
- Copyright© 2009 International Institute of Anticancer Research (Dr. John G. Delinassios), All rights reserved





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}