


Abstract
Background: High expression of tumor endothelial marker 7 (TEM7) is correlated with osteogenic sarcoma (OS) metastasis and poor survival of patients. The TEM7 gene produces four alternatively spliced transcripts with distinct functional domains; the expression pattern of these transcripts in OS is unknown. Materials and Methods: mRNA expression was assessed in 5 OS cell lines, 7 normal bone, and 9 OS tumor specimens by reverse transcriptase polymerase chain reaction. Results: All OS cell lines, 6/9 tumors but none of the bone specimens expressed mRNA of TEM7 secreted forms 1 and 2. A total of 3/5 OS cell lines, 8/9 of tumors and 4/7 of bone specimens expressed mRNA of the TEM7 intracellular form. One out of 5 cell lines, 2/7 tumors and none of the bone specimens expressed mRNA of the TEM7 membrane form. The secreted forms had 20-fold higher expression in metastatic (LM7) compared to non-metastatic (SAOS-2) cells. Conclusion: The mRNA of secreted and the membrane forms of TEM7 are preferentially expressed in OS.
Osteosarcoma (OS) is the most common primary bone tumor in children and adolescents. Five-year survival rates for localized disease have increased significantly over the past decades. In contrast, in metastatic disease, 5-year survival rates have reached a plateau and as yet have not further improved. A significant number of patients develop metastasis before diagnosis and have a poor prognosis, particularly because there is a lack of diagnostic molecular markers for OS. Therefore, identifying and evaluating potential diagnostic and prognostic markers deserves particular attention. However, only a few studies addressing this topic have been published (1-6). Except for the cytoskeleton membrane linker protein ezrin, there are only a few molecular markers which are clinically and prognostically relevant.
Tumor endothelial marker 7 (TEM7) was initially identified as the most abundantly expressed gene in a group of cell surface proteins in the vascular endothelium of human tumors (7). It is a gene associated with high-grade OS and metastasis (1). Elevated mRNA expression of TEM7 was reported in endothelial cells derived from tumors, and a role in normal tissues as well as in physiological and pathological processes such as tumor angiogenesis was described (7-10). TEM proteins are believed to be located on the cell surface. They have a single pass transmembrane protein with a conserved cytoplasmic tail (8, 10). TEM proteins contain a plexin-like as well as a weak nidogen-like domain, which may be involved in mediating protein—protein interactions (10).
Two major reasons dictate our interest in TEM7: the protein appears to be involved in OS metastasis (1), and the protein is believed to be expressed on the cell surface — we speculate that proteins which are expressed on the surface of tumor cells are better therapeutic targets than intracellular proteins simply because of easy accessibility of the protein to a drug. Recently, however, four alternatively spliced variants of TEM7 transcripts have been reported: a membrane-bound form (T-m), which contains a signal peptide and a transmembrane domain; two secreted forms (T-s1, T-s2) containing the signal peptide domains but lacking the transmembrane domain, and one intracellular form (T-i) that lacks both the signal peptide and the transmembrane domains. Furthermore, the secreted and the intracellular forms also lack the plexin and cortactin-binding domains (10). In addition, T-m lacks exon 10 and contains exon 14, which codes for the transmembrane domain. PCR analyses, comparisons with expressed sequence tags and genomic sequencing revealed that all four of these TEM7 variants are derived by alternative splicing (10). The existence of multiple variants adds a new dimension to the potential biological function of the molecule; we reasoned that the first step to uncover the role of TEM7 in OS metastasis would be to identify the variant(s) specifically expressed in OS. Consequently, we have evaluated expression of the four TEM7 transcripts in normal bone and OS cell lines, as well as OS tumor specimens. It appears that for various reasons, reverse transcriptase polymerase chain reaction is the only method that can be employed to assess expression of the alternatively spliced transcripts, which is what has been used in this study.
Primers used for RT-PCR of the TEM7 alternatively spliced transcripts. Sequences were obtained from the accession numbers AF279144 (for the membrane-specific TEM7 mRNA), AY704670 (for the intracellular form of Tem7 mRNA), AY704671 (for the S2 secreted form of TEM7 mRNA) and AY704672 (for the S1 secreted form of TEM7 mRNA).
Materials and Methods
Cell lines, tissue samples and reagents. FOB (immortalized normal human fetal osteoblast cell line), MG-63, SaOS-2, TE85 and 143B were from ATCC (Rockville, MD, USA). LM7 cells were obtained from Dr. E. Kleinerman (MD Anderson Cancer Center, Houston, TX, USA). Tissue specimens were procured from patients operated on for OS at our institution through approved IRB protocol. Normal bone cell lines were generated in our laboratory from surgical waste specimens (‘Normal bone’ here refers to bone samples from patients not having any tumor involving the skeleton. “Normal bone cell lines” refer to cells that grew when the above crushed and collagen-treated bone specimens were put into complete culture media). All cells were grown in antibiotic-free Dulbecco's modified Eagle's medium (Invitrogen, Carlsbad, CA, USA) and F12 (Invitrogen) (1:1) supplemented with 10% heat-inactivated fetal calf serum (FCS). Cells were cultured at 37°C in an atmosphere of 5% CO2 and 95% relative humidity.
Isolation of RNA. Total RNA was isolated from cells grown to 80% confluence using TRI reagent (Sigma-Aldrich, St. Louis, MO, USA) as recommended by the supplier. The RNA was quantified by measuring optical density at 260 and 280 nm in a UV-spectrophotometer (Beckman Instruments, Fullerton, CA, USA). The integrity of the RNA was assessed by standard agarose gel electrophoresis.
Reverse transcription and PCR. cDNA was reversely transcribed from 2 μg of total RNA with the Stratagene first-strand synthesizing system and random primers (Stratagene, La Jolla, CA, USA) according to the protocol supplied by the manufacturer. PCR primers (Table I) were synthesized at Mayo Molecular Core Facility. PCR was performed in a total volume of 20 μl containing 2 μl of cDNA (equivalent to a concentration of 5 ng/μl of RNA in the reverse transcription (RT) reaction), 2 μl of 10X PCR buffer, 2 μl of 10x dNTP mix, 1 μl of each primer (0.2 μM final), and 1 U of AmpliTaq polymerase (Applied Biosystems, Foster City, CA, USA). PCR was carried out in a thermocycler with initial denaturation at 94°C for 5 min followed by 30 cycles of denaturation at 94°C for 60 s, annealing at 65°C for 60 s and elongation at 72°C for 60 s. PCR was completed by further elongation at 72°C for 10 min. PCR products were analyzed by 2% agarose gel electrophoresis in the presence of ethidium-bromide (0.1‰). The identities of the PCR products were confirmed by direct sequencing.
Quantitative real-time PCR. Quantitative real-time PCR analysis of TEM7 transcripts was performed with a Light Cycler 2.0 instrument (Roche, Rotkreuz, Switzerland). The levels of individual transcripts were determined in cDNA aliquots equivalent to 0.4 μl of the reverse transcription reaction (from a total of 10 μl) of RNA isolated from ~20,000 80% confluent cells. The primers were identical to those used for semi-quantitative PCR. PCR was carried out according to the manufacturer's instructions (Roche) in a total volume of 20 μl containing 0.4 μl cDNA, 12 μl PCR grade water, primer at 4 μM concentration and 4 μl of Light Cycler Fast Start DNA Master Plus SYBR I ready-to-use “Hot Start” reaction mixture (Roche).
The reaction conditions were: pre-incubation at 95°C for 10 min, 50 cycles of denaturation at 95°C, primer-specific annealing (Table I) for 7 s, and elongation at 72°C for 7 s. Melting curves were recorded for 5 min from 65°C to 95°C increasing at a rate of 0.1°C/s. Experiments were performed in duplicates and the Cp values reflecting relative expression levels of selected genes were determined. Relative quantification was carried out using a calibrator (10 ng/μl) in the respective PCR reaction.
Results
The extremely high sequence identity among the four TEM7 splice variants poses a daunting task for designing PCR primers with the goal of obtaining variant-specific amplification products. To design primers, we first aligned the four cDNA sequences as published, and manually scanned for locations (sequences) specific for a variant. We failed to find sequences for any variant, which could be used to design a primer set (two primers) that would, theoretically, amplify a given variant only. Consequently, concessions were allowed to generate best possible variant-specific primer pairs. Primer sequences and other relevant information are presented in Table I.
Two primers Ti-14-26D and Ti-263-24U were designed for amplifying a segment from the T-i variant mRNA. The sequence of the primer Ti-263-24U is common to all the variants; however, that for the primer Ti-14-26D is specific for the intracellular variant. Consequently, specific amplification product was expected with this specific primer pair. Creating appropriate primers for the T-s1 variant posed the greatest challenge: Two primers Ts1-862-24D and Ts1-1090-24U were designed for amplifying a segment for this transcript. As presented in Table I, the first primer is common to all the variants, while the first 18 nucleotides of the second primer are common to T-s1, T-s2 and T-i variants but not to T-m (the sequence is absent in T-m). The last five nucleotides of this primer are also common to all the variants; however, the 19th nucleotide (a ‘C’) is specific for the T-s1 variant. We speculated that these primer pairs would produce amplification products for the T-s1 and T-i variants but not the other two variants. Two primers, Ts2-939-22D and Ts2-1094-24U, were designed for specific amplification of a segment from the T-s2 variant. While sequence of the first primer is common to all the variants, the sequence of the second primer is completely specific for the T-s2 variant and consequently a T-s2-specific PCR product was expected. A specific amplification product was also expected for the T-m variant because although the primer Tm-901-25D has a sequence common to all the variants and the first 19 nucleotides of the primer Tm-1090-23U is also common for all the variants, the last five nucleotides of the latter are specific for the T-m variant, consequently we expected a specific amplification product since the nucleotides at the 3′-end of a primer are among the most important determinants for specific annealing and elongation of DNA synthesis during PCR. It is important to note that even though a variant-specific primer pair may amplify the specific as well as segments from other variants, the identity of a specific product can be deduced from the length of the amplified DNA (see Table I).
We first evaluated expression of the four TEM7 transcript variants in five OS-derived cell lines and in FOB cells by semi-quantitative RT-PCR. Based on our animal model of spontaneous lung metastasis of OS (11), MG63 is a non-tumorigenic cell line, SAOS and TE85 are tumorigenic but poorly metastatic cell lines, and LM7 (a derivative of SAOS) and 143B (a derivative of TE85) are highly metastatic cell lines. LM7 and 143B cells have been observed in our laboratory to cause lung metastasis in 100% of animals (11).
Representative RT-PCR results are presented in Figure 1. The T-m variant had a high expression in SAOS cells only (panel A); the FOB cells also appeared to express the variant but at a very low level. The T-s1 variant was expressed in all cell lines including FOB (panel B), whereas the T-s2 variant is expressed in all 5 OS-derived cells but not in FOB (panel C). The T-i variant was expressed by FOB cells as well as by 3 out of 5 OS-derived cells (panel D).
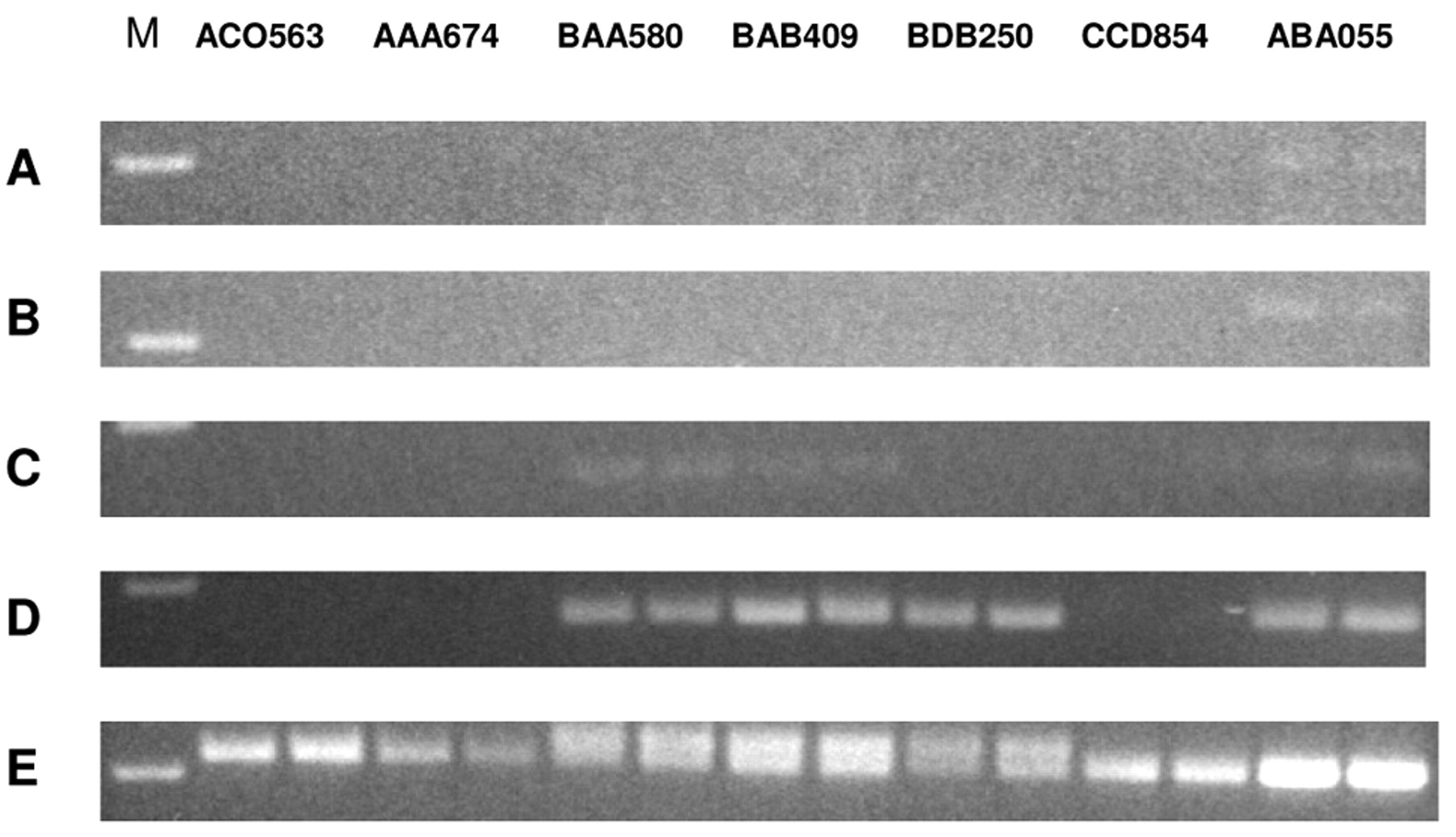
The pattern of expression of the four TEM7 alternatively spliced variants in normal bone, as determined by semi-quantitative RT-PCR, is presented in Figure 2. As can be seen, only the intracellular mRNA variant appeared to be expressed in bone (4 out of 7 samples). The T-m and T-s1 variants were expressed at a very low level in one sample only (ABA055), whereas low-level expression of the T-s2 variant was seen in 4/7 samples. Thus, the intracellular TEM7 variant mRNA appears to be the predominant form expressed in normal bone.
We also examined the expression pattern of the four TEM7 mRNA variants in 9 primary OS tumor specimens, and the results are presented in Figure 3. As can be seen, the T-m variant was expressed in 3 out of 9 tumor specimens (intense PCR-amplified product in JH21 and GS53, less intense product in SC14), whereas the T-s1 and T-s2 variants were expressed in 8 of 9 and in 7 out of 9 samples, respectively. The T-i variant mRNA appears to be expressed in all 9 tumor samples. Thus, except for the T-m variant, expression of the other TEM7 mRNA variants was frequent in these tumor samples. Note that for GAPDH, we routinely obtain a clean amplification product from cell lines, however, we sometimes obtained two amplified DNA bands from some whole tissue samples (normal or tumor tissues, containing cells of diverse types) — upon sequencing, one of these ‘doublet’ bands was found to originate from a GAPDH pseudogene (data not shown).
A summary of the RT-PCR results for all the TEM7 mRNA variants in OS-derived cell lines, normal bone specimens and OS tumors is presented in Table II. It can be inferred from these results that although expression of the T-m variant is infrequent, it is tumor specific. Expression of the T-i variant is not tumor specific since it was also expressed in 4 out of 7 bone samples. However, expression of the secreted forms appears to be tumor specific since they were not expressed in any of the normal bone samples tested.

Semi-quantitative RT-PCR to evaluate the expression level of the four alternatively spliced TEM7 transcripts in cell lines. A, T-m variant; B, T-s1 variant; C, T-s2 variant; D, T-1 variant; E, GAPDH mRNA. M, 100 bp DNA ladder. Cell line identifications are on top.
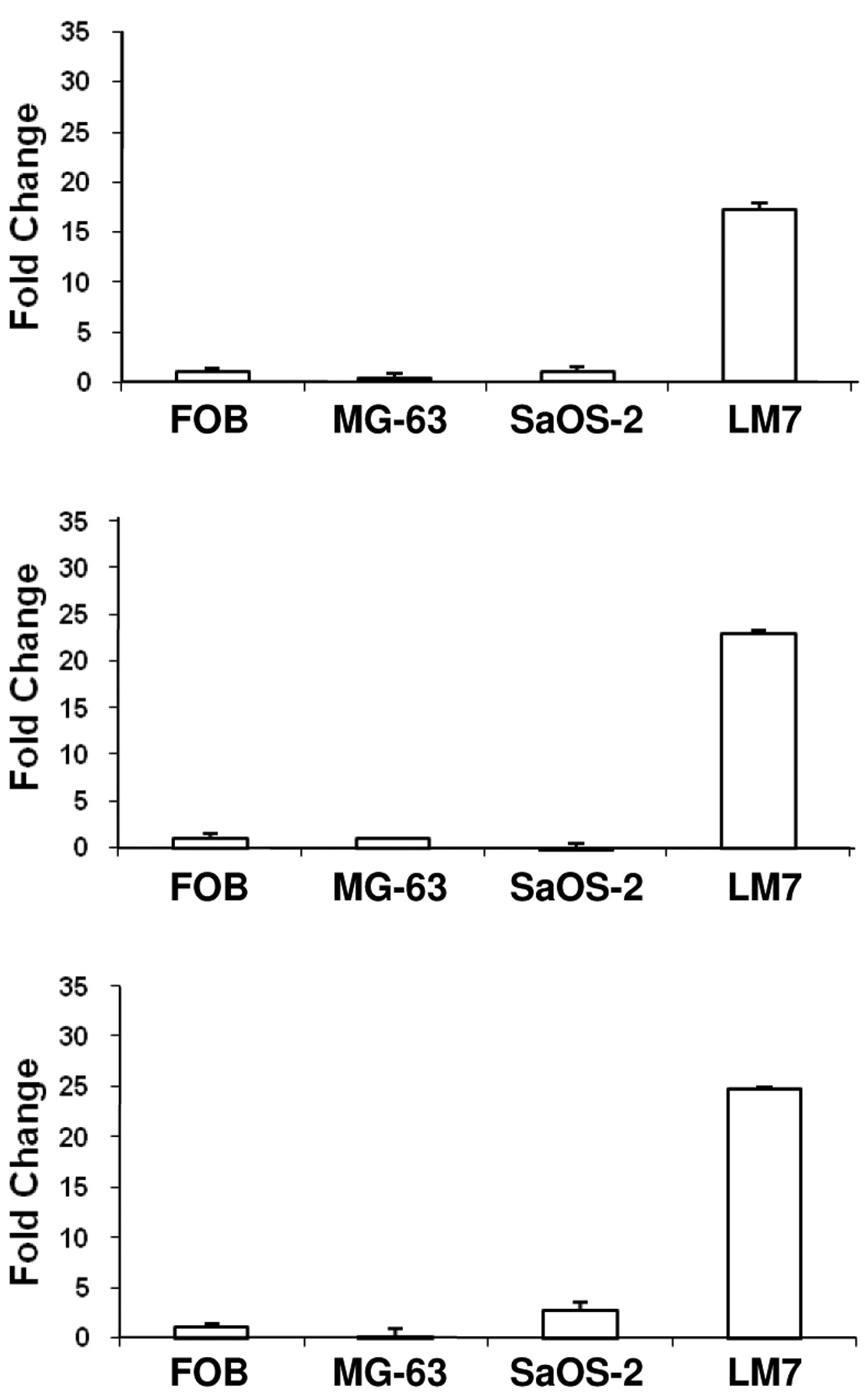
We have reported previously a significant association of TEM7 expression with OS metastasis. From results presented above, it appears that the T-s and/or the T-m forms of TEM7 may underlie this association. We performed quantitative PCR to compare the level of these three forms of TEM7 mRNA in non-metastatic, poorly metastatic and highly metastatic OS-derived cell lines; results from these experiments are presented in Figure 4. All three TEM7 mRNA variants were highly expressed in the metastatic LM7 cells compared to the poorly metastatic SAOS cells (>20-fold). The non-tumorigenic MG63 cells and the immortalized FOB cells had very little expression of the mRNAs. These results provide further support for a relationship between TEM7 expression and metastasis (1).
Discussion
The family of TEM proteins was initially identified as a result of a search specifically targeting the endothelium of tumor blood vessels as a new anticancer regimen (7). We recently showed a significant association between TEM7 expression level and metastasis of osteosarcoma (1). Recently, four alternatively spliced forms of TEM7 mRNA have been reported (10).
Since these forms are predicted to have different domains at the protein level, these variant TEM7 molecules could potentially create a diverse array of biological function. It is not known for any tumor type if all or some of these variants are tumor specific; consequently, our goal was to identify the variant(s), if any, specifically expressed in OS specimens. Our results strongly suggest that both the secreted and the membrane forms of TEM7 transcripts are specifically expressed in OS specimens, whereas the intracellular form appears to be common for both normal bone and OS cells. Our results further suggest that the membrane and the secreted forms of TEM7 are involved in the metastasis of OS. Thus, the TEM7 T-i transcript appears to be the ‘normal’ transcript; OS cells at some stage of development (probably during the course of metastasis) acquire the extra spliced forms of TEM7 mRNA. Consequently, we speculate that the OS cells, because of their possession of extra forms of TEM7 molecules acquire a novel functional phenotype, which, in turn, confers a survival advantage to the cancer cells.
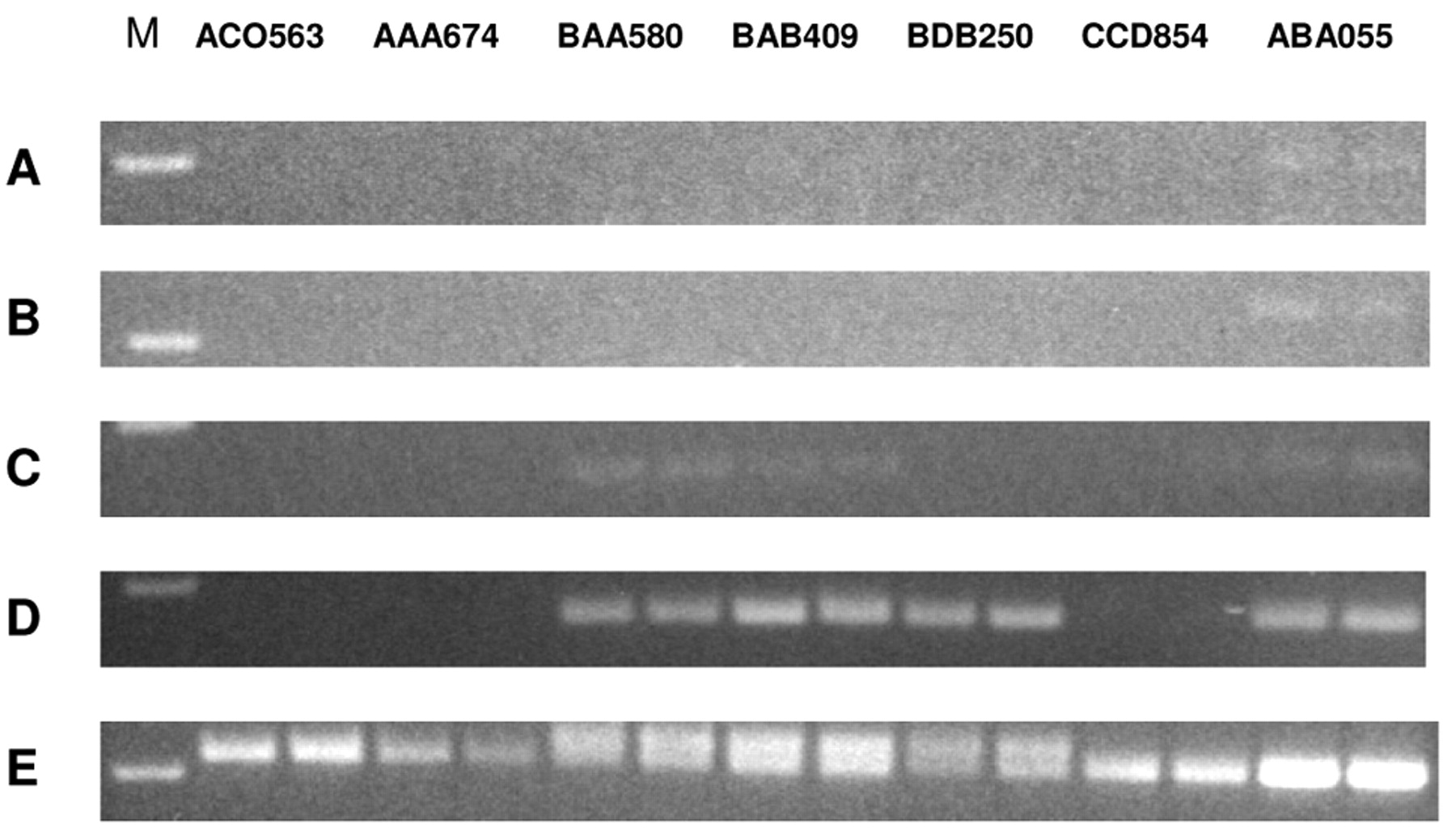
Semi-quantitative RT-PCR to evaluate the expression level of the four alternatively spliced TEM7 transcripts in cultured osteoblasts prepared in the laboratory from bone biopsy specimens. A, T-m variant; B, T-s1 variant; C, T-s2 variant; D, T-1 variant; E, GAPDH mRNA. M, 100 bp DNA ladder. Sample identifications are on top.

Semi-quantitative RT-PCR to evaluate the expression level of the four alternatively spliced TEM7 transcripts in OS tumor specimens. A, T-m variant; B, T-s1 variant; C, T-s2 variant; D, T-1 variant; E, GAPDH mRNA. M, 100 bp DNA ladder. Tumor specimen identifications are on top.
Summary of expression of TEM7 variants in various samples by RT-PCR.
The differential expression pattern of the TEM7 mRNA variants between normal bone and OS cells as reported herein provides a conceptual basis as to how these splice variants may carry out functions needed by the cancer cells. The T-m variant has been shown to contain a cortactin-binding site (10), which could mediate cell-cell interaction conducive to OS metastasis. In addition, the secreted forms of TEM7, which do not have a cortactin-binding site, may recognize different cell-surface and/or matrix proteins to activate signaling pathways favorable to OS cells. Thus, the membrane and the secreted forms of TEM7 together may contribute considerably to the proliferation and survival of OS cells. It would be interesting to evaluate if the expression pattern of TEM7 mRNA variants reported herein would be similar in cancer types other than OS.
In situ hybridization is a robust technique to evaluate mRNA expression in a specimen. However, it may not be applicable to assess the level of expression of the various TEM7 transcripts because it appears almost impossible to design probes specific for each of the variants. Moreover, due to size similarity, Northern blotting may not be applicable to distinguish the splice variants with confidence. Furthermore, since variant-specific antibodies are not available (and may be extremely difficult to generate), studies were not carried out on the expression pattern of the cognate proteins from the respective TEM7 mRNA variants. However, we have begun exploring the role of the alternatively spliced TEM7 mRNA variants in OS metastasis by employing siRNA. In conclusion, we present a first-ever study on the expression of TEM7 mRNA variants, and our results should provide a useful platform to further explore the potential role of TEM7 in osteosarcoma.

Quantitative RT-PCR to determine the expression level of T-m, T-s1 and T-s2 transcripts in OS cell lines. FOB, An immortalized human fetal osteoblast cell line; MG-63, a non-tumorigenic OS cell line; SAOS-2, a poorly metatstatic OS cell line; LM7, a highly metastatic derivative of SAOS-2.
Acknowledgements
This work was supported by a grant from the National Institute of Health (AR47974).
Footnotes
-
↵* Both authors contributed equally to this work.
- Received June 5, 2009.
- Revision received September 1, 2009.
- Accepted September 21, 2009.
- Copyright© 2009 International Institute of Anticancer Research (Dr. John G. Delinassios), All rights reserved





{kind=link}
{kind=link}
{kind=link}
{kind=link}